BIOLOGICAL SCIENCES
CHAPTER 1
INTRODUCTION: THEMES IN
THE STUDY OF LIFE
OUTLINE
I. Life’s Hierarchical Order
A. The living world is a hierarchy, with each level of biological structure building on the level below it
B. Each level of biological structure has emergent properties
C. Cells are an organism’s basic units of structure and function
D. The continuity of life is based on heritable information in the form of DNA E. Structure and function are correlated at all levels of biological organization F. Organisms are open systems that interact continuously with their environments G. Regulatory mechanisms ensure a dynamic balance in living systems
II. Evolution, Unity, and Diversity
A. Diversity and unity are the dual faces of life on Earth
B. Evolution is the core theme of biology
III. Science as a Process
A. Testable hypotheses are the hallmarks of the scientific process
B. Science and technology are functions of society
C. Biology is a multidisciplinary adventure
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Briefly describe unifying themes that pervade the science of biology. 2. Diagram the hierarchy of structural levels in biology.
3. Explain how the properties of life emerge from complex organization. 4. Describe seven emergent properties associated with life.
5. Distinguish between holism and reductionism.
6. Explain how technological breakthroughs contributed to the formulation of the cell theory and our current knowledge of the cell.
7. Distinguish between prokaryotic and eukaryotic cells.
8. Explain, in their own words, what is meant by "form fits function."
9. List the five kingdoms of life and distinguish among them.
10. Briefly describe how Charles Darwin's ideas contributed to the conceptual framework of biology.
11. Outline the scientific method.
12. Distinguish between inductive and deductive reasoning.
13. Explain how science and technology are interdependent.
2 Chapter 1 Introduction: Themes in the Study of Life
KEY TERMS
emergent property holism evolution control group population reductionism natural selection variable community prokaryotic scientific method experimental group ecosystem eukaryotic hypothesis deductive reasoning biome taxonomy inductive reasoning scientific theory biogenesis
LECTURE NOTES
Biology, the study of life, is a human endeavor resulting from an innate attraction to life in its diverse forms (E.O. Wilson's biophilia).
The science of biology is enormous in scope.
• It reaches across size scales from submicroscopic molecules to the global distribution of biological communities.
• It encompasses life over huge spans of time from contemporary organisms to ancestral life forms stretching back nearly four billion years.
As a science, biology is an ongoing process.
• As a result of new research methods developed over the past few decades, there has been an information explosion.
• Technological advances yield new information that may change the conceptual framework accepted by the majority of biologists.
With rapid information flow and new discoveries, biology is in a continuous state of flux. There are, however, enduring unifying themes that pervade the science of biology: • A hierarchy of organization
• The cellular basis of life
• Heritable information
• The correlation between structure and function
• The interaction of organisms with their environment
• Unity in diversity
• Evolution: the core theme
• Scientific process: the hypothetico-deductive method
I. Life’s Hierarchical Order
A. The living world is a hierarchy, with each level of biological structure building on the level below it
A characteristic of life is a high degree of order. Biological organization is based on a hierarchy of structural levels, with each level building on the levels below it.
Chapter 1 Introduction: Themes in the Study of Life 3
Atoms
Complex biological molecules
Subcellular organelles
are ordered into
Cells
In multicellular organisms similar cells are organised into
Tissues
Organs
Organ systems
Complex organism
There are levels of organization beyond the individual organism:
Population = Community = Ecosystem =
Biomes = Biosphere =
Localized group of organisms belonging to the same species
Populations of species living in the same area
An energy-processing system of community interactions that include abiotic environmental factors such as soil and water
Large scale communities classified by predominant vegetation type and distinctive combinations of plants and animals
The sum of all the planet's ecosystems
B. Each level of biological organization has emergent properties Emergent property = Property that emerges as a result of interactions between components.
• With each step upward in the biological hierarchy, new properties emerge that were not present at the simpler organizational levels.
• Life is difficult to define because it is associated with numerous emergent properties that reflect a hierarchy of structural organization.
Some of the emergent properties and processes associated with life are the following: 1. Order. Organisms are highly ordered, and other characteristics of life emerge from this complex organization.
4 Chapter 1 Introduction: Themes in the Study of Life
2. Reproduction. Organisms reproduce; life comes only from life (biogenesis). 3. Growth and Development. Heritable programs stored in DNA direct the species-specific pattern of growth and development.
4. Energy Utilization. Organisms take in and transform energy to do work, including the maintenance of their ordered state.
5. Response to Environment. Organisms respond to stimuli from their environment.
6. Homeostasis. Organisms regulate their internal environment to maintain a steady-state, even in the face of a fluctuating external environment.
7. Evolutionary Adaptation. Life evolves in response to interactions between organisms and their environment.
Because properties of life emerge from complex organization, it is impossible to fully explain a higher level of order by breaking it into its parts.
Holism = The principle that a higher level of order cannot be meaningfully explained by examining component parts in isolation.
• An organism is a living whole greater than the sum of its parts.
• For example, a cell dismantled to its chemical ingredients is no longer a cell. It is also difficult to analyze a complex process without taking it apart.
Reductionism = The principle that a complex system can be understood by studying its component parts.
• Has been a powerful strategy in biology
• Example: Watson and Crick deduced the role of DNA in inheritance by studying its molecular structure.
The study of biology balances the reductionist strategy with the goal of understanding how the parts of cells, organisms, and populations are functionally integrated.
C. Cells are an organism’s basic units of structure and function
The cell is an organism's basic unit of structure and function.
• Lowest level of structure capable of performing all activities of life.
• All organisms are composed of cells.
• May exist singly as unicellular organisms or as subunits of multicellular organisms.
The invention of the microscope led to the discovery of the cell and the formulation of the cell theory.
• Robert Hooke (1665) reported a description of his microscopic examination of cork. Hooke described tiny boxes which he called "cells" (really cell walls). The significance of this discovery was not recognized until 150 years later.
• Antonie van Leeuwenhok (1600's) used the microscope to observe living organisms such as microorganisms in pond water, blood cells, and animal sperm cells.
• Matthias Schleiden and Theodor Schwann (1839) reasoned from their own microscopic studies and those of others, that all living things are made of cells. This formed the basis for the cell theory.
• The cell theory has since been modified to include the idea that all cells come from preexisting cells.
Over the past 40 years, use of the electron microscope has revealed the complex ultrastructure of cells.
• Cells are bounded by plasma membranes that regulate passage of materials between the cell and its surroundings.
• All cells, at some stage, contain DNA.
Chapter 1 Introduction: Themes in the Study of Life 5
Based on structural organization, there are two major kinds of cells: prokaryotic and eukaryotic.
Prokaryotic cell = Cell lacking membrane-bound organelles and a membrane-enclosed nucleus.
• Found only in the archaebacteria and bacteria
• Generally much smaller than eukaryotic cells
• Contains DNA that is not separated from the rest of the cell, as there is no membrane-bound nucleus
• Lacks membrane-bound organelles
• Almost all have tough external walls
Eukaryotic cell = Cell with a membrane-enclosed nucleus and membrane-enclosed organelles.
• Found in protists, plants, fungi, and animals
• Subdivided by internal membranes into different functional compartments called organelles
• Contains DNA that is segregated from the rest of the cell. DNA is organized with proteins into chromosomes that are located within the nucleus, the largest organelle of most cells.
• Cytoplasm surrounds the nucleus and contains various organelles of different functions
• Some cells have a tough cell wall outside the plasma membrane (e.g., plant cells). Animal cells lack cell walls.
Though structurally different, eukaryotic and prokaryotic cells have many similarities, especially in their chemical processes.
D. The continuity of life is based on heritable information in the form of DNA
Biological instructions for an organism's complex structure and function are encoded in DNA.
• Each DNA molecule is made of four types of chemical building blocks called nucleotides.
• The linear sequence of these four nucleotides encode the precise information in a gene, the unit of inheritance from parent to offspring.
• An organism's complex structural organization is specified by an enormous amount of coded information.
Inheritance is based on:
• A complex mechanism for copying DNA.
• Passing the information encoded in DNA from parent to offspring. All forms of life use essentially the same genetic code.
• A particular nucleotide sequence provides the same information to one organism as it does to another.
• Differences among organisms reflect differences in nucleotide sequence.
E. Structure and function are correlated at all levels of biological organization
There is a relationship between an organism's structure and how it works. Form fits function.
• Biological structure gives clues about what it does and how it works. • Knowing a structure's function gives insights about its construction. • This correlation is apparent at many levels of biological organization.
6 Chapter 1 Introduction: Themes in the Study of Life
F. Organisms are open systems that interact continuously with their environments
Organisms interact with their environment, which includes other organisms as well as abiotic factors.
• Both organism and environment are affected by the interaction between them. • Ecosystem dynamics include two major processes:
1. Nutrient cycling
2. Energy flow (see Campbell, Figure 1.7)
G. Regulatory mechanisms ensure a dynamic balance in living systems Regulation of biological processes is critical for maintaining the ordered state of life. Many biological processes are self-regulating; that is, the product of a process regulates that process (= feedback regulation; see Campbell, Figure 1.8).
• Positive feedback speeds a process up
• Negative feedback slows a process down
Organisms and cells also use chemical mediators to help regulate processes.
II. Evolution, Unity, and Diversity
A. Diversity and unity are the dual faces of life on Earth
Biological diversity is enormous.
• Estimates of total diversity range from five million to over 30 million species. • About 1.5 million species have been identified and named, including approximately 260,000 plants, 50,000 vertebrates, and 750,000 insects.
To make this diversity more comprehensible, biologists classify species into categories. Taxonomy = Branch of biology concerned with naming and classifying organisms. • Taxonomic groups are ranked into a hierarchy from the most to least inclusive category: domain, kingdom, phylum, class, order, family, genus, species.
• A six-kingdom system recognizes two prokaryotic groups and divides the Monera into the Archaebacteria and Eubacteria.
• The kingdoms of life recognized in the traditional five-kingdom system are Monera, Protista, Plantae, Fungi, and Animalia (see Campbell, Figure 1.10). There is unity in the diversity of life forms at the lower levels of organization. Unity of life forms is evident in:
• A universal genetic code.
• Similar metabolic pathways (e.g., glycolysis).
• Similarities of cell structure (e.g., flagella of protozoans and mammalian sperm cells).
B. Evolution is the core theme of biology
Evolution is the one unifying biological theme.
• Life evolves. Species change over time and their history can be described as a branching tree of life.
• Species that are very similar share a common ancestor at a recent branch point on the phylogenetic tree.
• Less closely related organisms share a more ancient common ancestor.
Chapter 1 Introduction: Themes in the Study of Life 7
• All life is connected and can be traced back to primeval prokaryotes that existed more than three billion years ago.
In 1859, Charles Darwin published On the Origin of Species in which he made two major points:
1. Species change, and contemporary species arose from a succession of ancestors through a process of "descent with modification."
2. A mechanism of evolutionary change is natural selection.
Darwin synthesized the concept of natural selection based upon the following observations:
• Individuals in a population of any species vary in many inheritable traits. • Populations have the potential to produce more offspring than will survive or than the environment can support.
• Individuals with traits best suited to the environment leave a larger number of offspring, which increases the proportion of inheritable variations in the next generation. This differential reproductive success is what Darwin called natural selection.
Organisms' adaptations to their environments are the products of natural selection. • Natural selection does not create adaptations; it merely increases the frequency of inherited variants that arise by chance.
• Adaptations are the result of the editing process of natural selection. When exposed to specific environmental pressures, certain inheritable variations favor the reproductive success of some individuals over others.
Darwin proposed that cumulative changes in a population over long time spans could produce a new species from an ancestral one.
Descent with modification accounts for both the unity and diversity of life. • Similarities between two species may be a reflection of their descent from a common ancestor.
• Differences between species may be the result of natural selection modifying the ancestral equipment in different environmental contexts.
III. Science as a Process
A. Testable hypotheses are the hallmarks of the scientific process As the science of life, biology has the characteristics associated with science in general. Science is a way of knowing. It is a human endeavor that emerges from our curiosity about ourselves, the world, and the universe. Good scientists are people who:
• Ask questions about nature and believe those questions are answerable. • Are curious, observant, and passionate in their quest for discovery.
• Are creative, imaginative, and intuitive.
• Are generally skeptics.
Scientific method = Process which outlines a series of steps used to answer questions. • Is not a rigid procedure.
• Based on the conviction that natural phenomena have natural causes. • Requires evidence to logically solve problems.
The key ingredient of the scientific process is the hypothetico-deductive method, which is an approach to problem-solving that involves:
1. Asking a question and formulating a tentative answer or hypothesis by inductive reasoning.
2. Using deductive reasoning to make predictions from the hypothesis and then testing the validity of those predictions.
8 Chapter 1 Introduction: Themes in the Study of Life
Hypothesis = Educated guess proposed as a tentative answer to a specific question or problem.
Inductive reasoning = Making an inference from a set of specific observations to reach a general conclusion.
Deductive reasoning = Making an inference from general premises to specific consequences, which logically follow if the premises are true.
• Usually takes the form of If...then logic.
• In science, deductive reasoning usually involves predicting experimental results that are expected if the hypothesis is true.
Useful hypotheses have the following characteristics:
• Hypotheses are possible causes. Generalizations formed by induction are not necessarily hypotheses. Hypotheses should also be tentative explanations for observations or solutions to problems.
• Hypotheses reflect past experience with similar questions. Hypotheses are not just blind propositions, but are educated guesses based upon available evidence. • Multiple hypotheses should be proposed whenever possible. The disadvantage of operating under only one hypothesis is that it might restrict the search for evidence in support of this hypothesis; scientists might bias their search, as well as neglect to consider other possible solutions.
• Hypotheses must be testable via the hypothetico-deductive method. Predictions made from hypotheses must be testable by making observations or performing experiments. This limits the scope of questions that science can answer.
• Hypotheses can be eliminated, but not confirmed with absolute certainty. If repeated experiments consistently disprove the predictions, then we can assume that the hypothesis is false. However, if repeated experimentation supports the deductions, we can only assume that the hypothesis may be true; accurate predictions can be made from false hypotheses. The more deductions that are tested and supported, the more confident we can be that the hypothesis is true.
Another feature of the scientific process is the controlled experiment which includes control and experimental groups.
Control group = In a controlled experiment, the group in which all variables are held constant.
• Controls are a necessary basis for comparison with the experimental group, which has been exposed to a single treatment variable.
• Allows conclusions to be made about the effect of experimental manipulation. • Setting up the best controls is a key element of good experimental design. Variable = Condition of an experiment that is subject to change and that may influence an experiment's outcome.
Experimental group = In a controlled experiment, the group in which one factor or treatment is varied.
Science is an ongoing process that is a self-correcting way of knowing. Scientists: • Build on prior scientific knowledge.
• Try to replicate the observations and experiments of others to check on their conclusions.
Chapter 1 Introduction: Themes in the Study of Life 9
• Share information through publications, seminars, meetings, and personal communication.
What really advances science is not just an accumulation of facts, but a new concept that collectively explains observations that previously seemed to be unrelated. • Newton, Darwin, and Einstein stand out in the history of science because they synthesized ideas with great explanatory power.
• Scientific theories are comprehensive conceptual frameworks which are well supported by evidence and are widely accepted by the scientific community.
B. Science and technology are functions of society
Science and technology are interdependent.
• Technology extends our ability to observe and measure, which enables scientists to work on new questions that were previously unapproachable.
• Science, in turn, generates new information that makes technological inventions possible.
• Example: Watson and Crick's scientific discovery of DNA structure led t o further investigation that enhanced our understanding of DNA, the genetic code, and how to transplant foreign genes into microorganisms. The biotechnology industry has capitalized on this knowledge to produce valuable pharmaceutical products such as human insulin.
We have a love-hate relationship with technology.
• Technology has improved our standard of living.
• The consequence of using technology also includes the creation of new problems such as increased population growth, acid rain, deforestation, global warming, nuclear accidents, ozone holes, toxic wastes, and endangered species.
• Solutions to these problems have as much to do with politics, economics, culture and values as with science and technology.
A better understanding of nature must remain the goal of science. Scientists should: • Try to influence how technology is used.
• Help educate the public about the benefits and hazards of specific technologies. C. Biology is a multidisciplinary adventure
Biology is a multidisciplinary science that integrates concepts from chemistry, physics and mathematics. Biology also embraces aspects of humanities and the social sciences.
10 Chapter 1 Introduction: Themes in the Study of Life
REFERENCES
Campbell, N. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Moore, J.A. "Science as a Way of Knowing–Evolutionary Biology." American Zoologist, 24(2): 470-475, 1980.
CHAPTER 2
THE CHEMICAL
CONTEXT OF LIFE
OUTLINE
I. Chemical Elements and Compounds
A. Matter consists of chemical elements in pure form and in combinations called compounds
B. Life requires about 25 chemical elements
II. Atoms and Molecules
A. Atomic structure determines the behavior of an element
B. Atoms combine by chemical bonding to form molecules
C. Weak chemical bonds play important roles in the chemistry of life
D. A molecule’s biological function is related to its shape
E. Chemical reactions make and break chemical bonds
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Define element and compound.
2. State four elements essential to life that make up 96% of living matter. 3. Describe the structure of an atom.
4. Define and distinguish among atomic number, mass number, atomic weight, and valence.
5. Given the atomic number and mass number of an atom, determine the number of neutrons.
6. Explain why radioisotopes are important to biologists.
7. Explain how electron configuration influences the chemical behavior of an atom. 8. Explain the octet rule and predict how many bonds an atom might form. 9. Explain why the noble gases are so unreactive.
10. Define electronegativity and explain how it influences the formation of chemical bonds.
11. Distinguish among nonpolar covalent, polar covalent and ionic bonds.
12. Describe the formation of a hydrogen bond and explain how it differs from a covalent or ionic bond.
13. Explain why weak bonds are important to living organisms.
14. Describe how the relative concentrations of reactants and products affect a chemical reaction.
12 Unit I The Chemistry of Life
KEY TERMS
matter atomic weight valence electron polar covalent bond element isotope valence shell ion trace element radioactive isotope chemical bond cation atom energy covalent bond anion neutron potential energy molecule ionic bond proton energy level structural formula hydrogen bond electron energy molecular formula chemical reactions atomic nucleus potential energy double covalent bond reactants dalton energy level valence products atomic number electron shell electronegativity chemical equilibrium mass number orbital nonpolar covalent bond
LECTURE NOTES
I. Chemical Elements and Compounds
A. Matter consists of chemical elements in pure form and in combinations called compounds
Chemistry is fundamental to an understanding of life, because living organisms are made of matter.
Matter = Anything that takes up space and has mass.
Mass = A measure of the amount of matter an object contains.
B. Life requires about 25 chemical elements
Element = A substance that cannot be broken down into other substances by chemical reactions.
• All matter is made of elements.
• There are 92 naturally occurring elements.
• They are designated by a symbol of one or two letters.
About 25 of the 92 naturally occurring elements are essential to life. Biologically important elements include:
C = carbon
O = oxygen make up 96% of living matter
H = hydrogen
N = nitrogen
Chapter 2 The Chemical Context of Life 13
Ca = calcium
P = phosphorus
K = potassium
S = sulfur make up remaining 4% of an organism's weight Na = sodium
Cl = chlorine
Mg = magnesium
Trace elements
Trace element = Element required by an organism in extremely minute quantities. • Though required by organisms in small quantity, they are indispensable for life • Examples: B, Cr, Co, Cu, F, I, Fe, Mn, Mo, Se, Si, Sn, V and Zn
Elements can exist in combinations called compounds.
• Compound = A pure substance composed of two or more elements combined in a fixed ratio.
• Example: NaCl (sodium chloride)
• Has unique emergent properties beyond those of its combined elements (Na and Cl have very different properties from NaCl). See Campbell, Figure 2.2.
II. Atoms and Molecules
A. Atomic structure determines the behavior of an element
Atom = Smallest possible unit of matter that retains the physical and chemical properties of its element.
• Atoms of the same element share similar chemical properties.
• Atoms are made up of subatomic particles.
1. Subatomic particles
The three most stable subatomic particles are:
1. Neutrons [no charge (neutral)].
2. Protons [+1 electrostatic charge].
3. Electrons [-1 electrostatic charge].
NEUTRON PROTON ELECTRON
No charge +1 charge -1 charge
Found together in a dense core called the nucleus (positively charged because of protons)
Orbits around nucleus (held by electrostatic attraction to positively charged nucleus)
1.009 dalton 1.007 dalton 1/2000 dalton
Masses of both are about the same (about 1 dalton) Mass is so small, usually not used to calculate
atomic mass
NOTE: The dalton is a unit used to express mass at the atomic level. One dalton (d) is equal to 1.67 x 10-24 g.
If an atom is electrically neutral, the number of protons equals the number of electrons, which yields an electrostatically balanced charge.
14 Unit I The Chemistry of Life
2. Atomic number and atomic weight
Atomic number = Number of protons in an atom of a particular element.
• All atoms of an element have the same atomic number.
• Written as a subscript to the left of the element's symbol (e.g., 11Na)
• In a neutral atom, # protons = # electrons.
Mass number = Number of protons and neutrons in an atom.
• Written as a superscript to left of an element's symbol (e.g., 23Na)
• Is approximate mass of the whole atom, since the mass of a proton and the mass of a neutron are both about 1 dalton
• Can deduce the number of neutrons by subtracting atomic number from mass number
• Number of neutrons can vary in an element, but number of protons is constant
• Is not the same as an element's atomic weight, which is the weighted mean of the masses of an element's constituent isotopes
Examples:
(Mass #) 23
(Atomic #) 11Na # of electrons
# of protons
# of neutrons
12
6C # of electrons
# of protons
# of neutrons
3. Isotopes
Isotopes = Atoms of an element that have the same atomic number but different mass number.
• They have the same number of protons, but a different number of neutrons. • Under natural conditions, elements occur as mixtures of isotopes.
• Different isotopes of the same element react chemically in same way. • Some isotopes are radioactive.
Radioactive isotope = Unstable isotope in which the nucleus spontaneously decays, emitting subatomic particles and/or energy as radioactivity.
• Loss of nuclear particles may transform one element to another
(e.g., 146C 147N).
• Has a fixed half life.
• Half life = Time for 50% of radioactive atoms in a sample to decay.
Biological applications of radioactive isotopes include:
a. Dating geological strata and fossils
Chapter 2 The Chemical Context of Life 15
• Radioactive decay is at a fixed rate.
• By comparing the ratio of radioactive and stable isotopes in a fossil with the ratio of isotopes in living organisms, one can estimate the age of a fossil.
• The ratio of 14C to 12C is frequently used to date fossils less than 50,000 years old.
b. Radioactive tracers
• Chemicals labelled with radioactive isotopes are used to trace the steps of a biochemical reaction or to determine the location of a particular substance within an organism (see Campbell, p. XX, Methods: The Use of Radioactive Tracers in Biology).
• Radioactive isotopes are useful as biochemical tracers because they chemically react like the stable isotopes and are easily detected at low concentrations.
• Isotopes of P, N, and H were used to determine DNA structure. • Used to diagnose disease (e.g., PET scanner)
• Because radioactivity can damage cell molecules, radioactive isotopes can also be hazardous
c. Treatment of cancer
• e.g., radioactive cobalt
4. The energy levels of electrons
Electrons = Light negatively charged particles that orbit around nucleus. • Equal in mass and charge
• Are the only stable subatomic particles directly involved in chemical reactions
• Have potential energy because of their position relative to the positively charged nucleus
Energy = Ability to do work
Potential energy = Energy that matter stores because of its position or location. • There is a natural tendency for matter to move to the lowest state of potential energy.
• Potential energy of electrons is not infinitely divisible, but exists only in discrete amounts called quanta.
• Different fixed potential energy states for electrons are called energy levels or electron shells (see Campbell, Figure 2.7).
• Electrons with lowest potential energy are in energy levels closest to the nucleus.
• Electrons with greater energy are in energy levels further from nucleus. Electrons may move from one energy level to another. In the process, they gain or lose energy equal to the difference in potential energy between the old and new energy level.
16 Unit I The Chemistry of Life
5. Electron orbitals
Orbital = Three-dimensional space where an electron will most likely be found 90% of the time (see Campbell, Figure 2.8).
• Viewed as a three-dimensional probability cloud (a statistical concept) • No more than two electrons can occupy same orbital.
First energy level:
• Has one spherical s orbital (1s orbital)
• Holds a maximum of two electrons
Second energy level
• Holds a maximum of eight electrons
• One spherical s orbital (2s orbital)
• Three dumbbell-shaped p orbitals each oriented at right angles to the other two (2px, 2py, 2pz orbitals)
Higher energy levels:
• Contain s and p orbitals
• Contain additional orbitals with more complex shapes
6. Electron configuration and chemical properties
An atom’s electron configuration determines its chemical behavior.
• Electron configuration = Distribution of electrons in an atom's electron shells
The first 18 elements of a periodic chart are arranged sequentially by atomic number into three rows (periods). In reference to these representative elements, note the following:
• Outermost shell of these atoms never have more than four orbitals (one s and three p) or eight electrons.
• Electrons must first occupy lower electron shells before the higher shells can be occupied. (This is a reflection of the natural tendency for matter to move to the lowest possible state of potential energy—the most stable state.)
• Electrons are added to each of the p orbitals singly, before they can be paired.
• If an atom does not have enough electrons to fill all shells, the outer shell will be the only one partially filled. Example: O2 with a total of eight electrons:
Chapter 2 The Chemical Context of Life 17
OXYGEN
8O
Two electrons have the 1s orbital of the first
electron shell.
First two electrons in the second shell are
both in the 2s orbital.
Next three electrons each have a p orbital
(2px, 2py, 2pz).
Eighth electron is paired in the 2px orbital. 1s2
2s 2px 2py 2pz
2 2 1 1
Chemical properties of an atom depend upon the number of valence electrons. • Valence electrons = Electrons in the outermost energy shell (valence shell). Octet rule = Rule that a valence shell is complete when it contains eight electrons (except H and He).
• An atom with a complete valence shell is unreactive or inert.
• Noble elements (e.g., helium, argon, and neon) have filled outer shells in their elemental state and are thus inert.
• An atom with an incomplete valence shell is chemically reactive (tends to form chemical bonds until it has eight electrons to fill the valence shell). • Atoms with the same number of valence electrons show similar chemical behavior.
NOTE: The consequence of this unifying chemical principle is that the valence electrons are responsible for the atom's bonding capacity. This rule applies to most of the representative elements, but not all.
B. Atoms combine by chemical bonding to form molecules
Atoms with incomplete valence shells tend to fill those shells by interacting with other atoms. These interactions of electrons among atoms may allow atoms to form chemical bonds.
• Chemical bonds = Attractions that hold molecules together
Molecules = Two or more atoms held together by chemical bonds.
1. Covalent bonds
Covalent bond = Chemical bond between atoms formed by sharing a pair of valence electrons.
• Strong chemical bond
• Example: molecular hydrogen (H2); when two hydrogen atoms come close H2 H H H-H
18 Unit I The Chemistry of Life
enough for their 1s orbitals to overlap, they share electrons, thus completing the valence shell of each atom.
Structural formula = Formula which represents the atoms and bonding within a molecule (e.g., H-H). The line represents a shared pair of electrons.
Molecular formula = Formula which indicates the number and type of atoms (e.g., H2).
Single covalent bond = Bond between atoms formed by sharing a single pair of valence electrons.
• Atoms may freely rotate around the axis of the bond.
Double covalent bond = Bond formed when atoms share two pairs of valence electrons (e.g., O2).
O2 O O O=O
Molecules = Two or more atoms held together by chemical bonds.
Triple covalent bond = Bond formed when atoms share three pairs of valence electrons (e.g., N2 or NºN).
NOTE: Double and triple covalent bonds are rigid and do not allow rotation. Valence = Bonding capacity of an atom which is the number of covalent bonds that must be formed to complete the outer electron shell.
• Valences of some common elements: hydrogen = 1, oxygen = 2, nitrogen = 3, carbon = 4, phosphorus = 3 (sometimes 5 as in biologically important compounds, e.g., ATP), sulfur = 2.
Compound = A pure substance composed of two or more elements combined in a fixed ratio.
• Example: water (H2O), methane (CH4)
• Note that two hydrogens are necessary to complete the valence shell of oxygen in water, and four hydrogens are necessary for carbon to complete the valence shell in methane.
Chapter 2 The Chemical Context of Life 19
2. Nonpolar and polar covalent bonds
Electronegativity = Atom’s ability to attract and hold electrons.
• The more electronegative an atom, the more strongly it attracts shared electrons.
• Scale determined by Linus Pauling:
O = 3.5
N = 3.0
S and C = 2.5
P and H = 2.1
Nonpolar covalent bond = Covalent bond formed by an equal sharing of electrons between atoms.
• Occurs when electronegativity of both atoms is about the same (e.g., CH4) • Molecules made of one element usually have nonpolar covalent bonds (e.g., H2, O2, Cl2, N2).
Polar covalent bond = Covalent bond formed by an unequal sharing of electrons between atoms.
• Occurs when the atoms involved have

different electronegativities.
• Shared electrons spend more time
around the more electronegative atom.
• In H2O, for example, the oxygen is
strongly electronegative, so negatively
charged electrons spend more time
around the oxygen than the hydrogens.
This causes the oxygen atom to have a
slight negative charge and the
hydrogens to have a slight positive
charge (see also Campbell, Figure 2.11).
3. Ionic bonds
Ion = Charged atom or molecule.
Anion = An atom that has gained one or more electrons from another atom and has become negatively charged; a negatively charged ion.
Cation = An atom that has lost one or more electrons and has become positively charged; a positively charged ion.
Ionic bond = Bond formed by the electrostatic attraction after the complete transfer of an electron from a donor atom to an acceptor.
• The acceptor atom attracts the electrons because it is much more electronegative than the donor atom.
• Are strong bonds in crystals, but are fragile bonds in water; salt crystals will readily dissolve in water and dissociate into ions.
• Ionic compounds are called salts (e.g., NaCl or table salt) (see Campbell, Figure 2.13).
NOTE: The difference in electronegativity between interacting atoms determines if electrons are shared equally (nonpolar covalent), shared unequally (polar covalent), gained or lost (ionic bond). Nonpolar covalent bonds and ionic bonds are two extremes of a continuum from interacting atoms with similar
electronegativities to interacting atoms with very different electronegativities.
20 Unit I The Chemistry of Life
C. Weak chemical bonds play important roles in the chemistry of life Biologically important weak bonds include the following:
• Hydrogen bonds, ionic bonds in aqueous solutions, and other weak forces such as Van der Waals and hydrophobic interactions
• Make chemical signaling possible in living organisms because they are only temporary associations. Signal molecules can briefly and reversibly bind t o receptor molecules on a cell, causing a short-lived response.
• Can form between molecules or between different parts of a single large molecule.
• Help stabilize the three-dimensional shape of large molecules (e.g., DNA and proteins).
1. Hydrogen bonds
Hydrogen bond = Bond formed by the charge attraction when a hydrogen atom covalently bonded to one electronegative atom is attracted to another
electronegative atom.
• Weak attractive force that is about
20 times easier to break than a covalent bond
• Is a charge attraction between oppositely charged portions of polar
HO
H
Electronegative
molecules
• Can occur between a hydrogen that has a slight positive charge when covalently bonded to an atom with high electronegativity (usually O and N)
• Example: NH3 in H2O (see Campbell, Figure 2.14)
2. Van der Waals interactions
Hydrogen atoms bond
N
H
HH
Weak interactions that occur between atoms and molecules that are very close together and result from charge asymetry in electron clouds.
D. A molecule’s biological function is related to its shape
A molecule has a charasteric size and shape.
The function of many molecules depends upon their shape
• Molecules with only two atoms are linear.
• Molecules with more than two atoms have more complex shapes.
When an atom forms covalent bonds, orbitals in the valence shell rearrange into the most stable configuration. To illustrate, consider atoms with valence electrons in the s and three p orbitals:
• The s and three p orbitals hybridize into four new orbitals.
• The new orbitals are teardrop shaped, extend from the nucleus and spread out as far apart as possible.
• Example: If outer tips of orbitals in methane (CH4) are connected by imaginary lines, the new molecule has a tetrahedral shape with C at the center (see Campbell, Figure 2.15).
Chapter 2 The Chemical Context of Life 21
E. Chemical reactions make and break chemical bonds
Chemical reactions = process of making and breaking chemical bonds leading to changes in the composition of matter.
• Process where reactants undergo changes into products.
• Matter is conserved, so all reactant atoms are only rearranged to form products. • Some reactions go to completion (all reactants converted to products), but most reactions are reversible. For example:
3H2 + N2 2NH3
• The relative concentration of reactants and products affects reaction rate (the higher the concentration, the greater probability of reaction).
Chemical equilibrium = Equilibrium established when the rate of forward reaction equals the rate of the reverse reaction.
• Is a dynamic equilibrium with reactions continuing in both directions
• Relative concentrations of reactants and products stop changing.
REFERENCES
Atkins, P.W. Atoms, Electrons and Change. W.H. Freeman and Company, 1991. Campbell, N., et al. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Weinberg, S. The Discovery of Subatomic Particles. New York, San Francisco: W.H. Freeman and Company, 1983.
Brown, T.L., H. E. Le May, Jr., and B. Bursten. Chemistry: The Central Science. 7th ed. Upper Saddle River, New Jersey: Prentice Hall, 1997.
CHAPTER 3
WATER AND THE FITNESS
OF THE ENVIRONMENT
OUTLINE
I. Water’s Polarity and Its Effects
A. The polarity of water molecules results in hydrogen bonding
B. Organisms depend on the cohesion of water molecules
C. Water moderates temperatures on Earth
D. Oceans and lakes don’t freeze solid because ice floats
E. Water is the solvent of life
II. The Dissociation of Water
A. Organisms are sensitive to changes in pH
III. Acid Precipitation Threatens the Fitness of the Environment
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Describe how water contributes to the fitness of the environment to support life. 2. Describe the structure and geometry of a water molecule, and explain what properties emerge as a result of this structure.
3. Explain the relationship between the polar nature of water and its ability to form hydrogen bonds.
4. List five characteristics of water that are emergent properties resulting from hydrogen bonding.
5. Describe the biological significance of the cohesiveness of water.
6. Distinguish between heat and temperature.
7. Explain how water's high specific heat, high heat of vaporization and expansion upon freezing affect both aquatic and terrestrial ecosystems.
8. Explain how the polarity of the water molecule makes it a versatile solvent. 9. Define molarity and list some advantages of measuring substances in moles. 10. Write the equation for the dissociation of water, and explain what is actually transferred from one molecule to another.
11. Explain the basis for the pH scale.
12. Explain how acids and bases directly or indirectly affect the hydrogen ion concentration of a solution.
13. Using the bicarbonate buffer system as an example, explain how buffers work.
24 Unit I The Chemistry of Life
14. Describe the causes of acid precipitation, and explain how it adversely affects the fitness of the environment.
KEY TERMS
polar molecule Celsius scale solute hydrogen ion cohesion calorie solvent molarity adhesion kilocalorie aqueous solution hydroxide ion surface tension joule hydrophilic acid kinetic energy specific heat hydrophobic base heat evaporative cooling mole pH scale temperature solution molecular weight buffer acid precipitation
LECTURE NOTES
Water contributes to the fitness of the environment to support life.
• Life on earth probably evolved in water.
• Living cells are 70%-95% H2O.
• Water covers about 3/4 of the earth.
• In nature, water naturally exists in all three physical states of matter—solid, liquid and gas.
Water's extraordinary properties are emergent properties resulting from water's structure and molecular interactions.
I. Water’s Polarity and Its Effects
A. The polarity of water molecules results in hydrogen bonding
Water is a polar molecule. Its polar bonds and asymmetrical shape give water molecules opposite charges on opposite sides.
• Four valence orbitals of O point t o corners of a tetrahedron.
• Two corners are orbitals with unshared pairs of electrons and weak negative charge.
• Two corners are occupied by H atoms which are in polar covalent bonds with O. Oxygen is so electronegative, that shared electrons spend more time around the O causing a weak positive charge near H's.
Hydrogen bonding orders water into a higher level of structural organization.
• The polar molecules of water are held together by hydrogen bonds.
• Positively charged H of one molecule is attracted to the negatively charged O of another water molecule.
Unbonded electron pairs
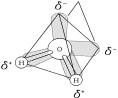
H
• Each water molecule can form a
O
H
maximum of four hydrogen bonds
with neighboring water molecules.
Chapter 3 Water and the Fitness of the Environment 25
Water has extraordinary properties that emerge as a consequence of its polarity and hydrogen-bonding. Some of these properties are that water:
• has cohesive behavior
• resists changes in temperature
• has a high heat of vaporization and cools surfaces as it evaporates • expands when it freezes
• is a versatile solvent
B. Organisms depend on the cohesion of water molecules.
Cohesion = Phenomenon of a substance being held together by hydrogen bonds. • Though hydrogen bonds are transient, enough water molecules are hydrogen bonded at any given time to give water more structure than other liquids. • Contributes to upward water transport in plants by holding the water column together. Adhesion of water to vessel walls counteracts the downward pull of gravity.
Surface tension = Measure of how difficult it is to stretch or break the surface of a liquid.
• Water has a greater surface tension than most liquids; function of the fact that at the air/H2O interface, surface water molecules are hydrogen bonded to each other and to the water molecules below.
• Causes H2O to bead (shape with smallest area to volume ratio and allows maximum hydrogen bonding).
C. Water moderates temperatures on Earth
1. Heat and temperature
Kinetic energy = The energy of motion.
Heat = Total kinetic energy due to molecular motion in a body of matter. Temperature = Measure of heat intensity due to the average kinetic energy of molecules in a body of matter.
Calorie (cal) = Amount of heat it takes to raise the temperature of one gram of water by one degree Celsius. Conversely, one calorie is the amount of heat that one gram of water releases when it cools down by one degree Celsius. NOTE: The “calories” on food packages are actually kilocalories (kcal).
Kilocalorie (kcal or Cal) = Amount of heat required to raise the temperature of one kilogram of water by one degree Celsius (1000 cal).
2. Water’s high specific heat
Water has a high specific heat, which means that it resists temperature changes when it absorbs or releases heat.
Specific heat = Amount of heat that must be absorbed or lost for one gram of a substance to change its temperature by one degree Celsius.
Specific heat of water = One calorie per gram per degree Celsius (1 cal/g/°C).
26 Unit I The Chemistry of Life
• As a result of hydrogen bonding among water molecules, it takes a relatively large heat loss or gain for each 1°C change in temperature.
• Hydrogen bonds must absorb heat to break, and they release heat when they form.
• Much absorbed heat energy is used to disrupt hydrogen bonds before water molecules can move faster (increase temperature).
A large body of water can act as a heat sink, absorbing heat from sunlight during the day and summer (while warming only a few degrees) and releasing heat during the night and winter as the water gradually cools. As a result:
• Water, which covers three-fourths of the planet, keeps temperature fluctuations within a range suitable for life.
• Coastal areas have milder climates than inland.
• The marine environment has a relatively stable temperature.
3. Evaporative cooling
Vaporization (evaporation) = transformation from liquid to a gas.
• Molecules with enough kinetic energy to overcome the mutual attraction of molecules in a liquid, can escape into the air.
Heat of vaporization = Quantity of heat a liquid must absorb for 1 g to be converted to the gaseous state.
• For water molecules to evaporate, hydrogen bonds must be broken which requires heat energy.
• Water has a relatively high heat of vaporization at the boiling point
(540 cal/g or 2260 J/g; Joule = 0.239 cal).
Evaporative cooling = Cooling of a liquid's surface when a liquid evaporates (see Campbell, Figure 3.4).
• The surface molecules with the highest kinetic energy are most likely to escape into gaseous form; the average kinetic energy of the remaining surface molecules is thus lower.
Water's high heat of vaporization:
• Moderates the Earth's climate.
• Solar heat absorbed by tropical seas dissipates when surface water evaporates (evaporative cooling).
• As moist tropical air moves poleward, water vapor releases heat as it condenses into rain.
• Stabilizes temperature in aquatic ecosystems (evaporative cooling).
• Helps organisms from overheating by evaporative cooling.
D. Oceans and lakes don’t freeze solid because ice floats
Because of hydrogen bonding, water is less dense as a solid than it is as a liquid. Consequently, ice floats.
• Water is densest at 4°C.
• Water contracts as it cools to 4°C.
• As water cools from 4°C to freezing (0°C), it expands and becomes less dense than liquid water (ice floats).
• When water begins to freeze, the molecules do not have enough kinetic energy to break hydrogen bonds.
• As the crystalline lattice forms, each water molecule forms a maximum of four hydrogen bonds, which keeps water molecules further apart than they would be in the liquid state; see Campbell, Figure 3.5.
Chapter 3 Water and the Fitness of the Environment 27
Expansion of water contributes to the fitness of the environment for life: • Prevents deep bodies of water from freezing solid from the bottom up. • Since ice is less dense, it forms on the surface first. As water freezes it releases heat to the water below and insulates it.
• Makes the transitions between seasons less abrupt. As water freezes, hydrogen bonds form releasing heat. As ice melts, hydrogen bonds break absorbing heat.
E. Water is the solvent of life
Solution = A liquid that is a completely homogenous mixture of two or more substances.
Solvent = Dissolving agent of a solution.
Solute = Substance dissolved in a solution.
Aqueous solution = Solution in which water is the solvent.
Water is a versatile solvent owing to the polarity of the water molecule.
Hydrophilic
{Ionic compounds dissolve in water (see Campbell, Figure
3.8).
• Charged regions of polar water molecules have an electrical attraction to charged ions.
• Water surrounds individual ions, separating and shielding them from one another.
Polar compounds in general, are water-soluble.
• Charged regions of polar water molecules have an affinity for oppositely charged regions of other polar
molecules.
Hydrophobic {Nonpolar compounds (which have symmetric distribution in charge) are NOT water-soluble.
1. Hydrophilic and hydrophobic substances
Ionic and polar substances are hydrophilic, but nonpolar compounds are hydrophobic.
Hydrophilic = (Hydro = water; philo = loving); property of having an affinity for water.
• Some large hydrophilic molecules can absorb water without dissolving.
Hydrophobic = (Hydro = water; phobos = fearing); property of not having an affinity for water, and thus, not being water-soluble.
2. Solute concentration in aqueous solutions
Most biochemical reactions involve solutes dissolved in water. There are two important quantitative properties of aqueous solutions: solute concentration and pH.
Molecular weight = Sum of the weight of all atoms in a molecule (expressed in daltons).
Mole = Amount of a substance that has a mass in grams numerically equivalent to its molecular weight in daltons.
28 Unit I The Chemistry of Life
For example, to determine a mole of sucrose (C12H22O11 ):
• Calculate molecular weight:
C = 12 dal 12 dal ⋅ 12 = 144 dal
H = 1 dal 1 dal ⋅ 22 = 22 dal
O = 16 dal 16 dal ⋅ 11 = 176 dal
342 dal
• Express it in grams (342 g).
Molarity = Number of moles of solute per liter of solution
• To make a 1M sucrose solution, weigh out 342 g of sucrose and add water up to 1L.
Advantage of measuring in moles:
• Rescales weighing of single molecules in daltons to grams, which is more practical for laboratory use.
• A mole of one substance has the same number of molecules as a mole of any other substance (6.02 ⋅ 1023 ; Avogadro's number).
• Allows one to combine substances in fixed ratios of molecules.
II. The Dissociation of Water
Occasionally, the hydrogen atom that is shared in a hydrogen bond between two water molecules, shifts from the oxygen atom to which it is covalently bonded to the unshared orbitals of the oxygen atom to which it is hydrogen bonded.
• Only a hydrogen ion (proton with a +1 charge) is actually transferred.
• Transferred proton binds to an unshared orbital of the second water molecule creating a hydronium ion (H3O+).
• Water molecule that lost a proton has a net negative charge and is called a hydroxide ion (OH-).
H2O + H2O H3O+ + OH-
• By convention, ionization of H2O is expressed as the dissociation into H+ and OH-.
H2O H+ + OH-
• Reaction is reversible.
• At equilibrium, most of the H2O is not ionized.
A. Organisms are sensitive to changes in pH
1. Acids and bases
At equilibrium in pure water at 25°C:
• Number of H+ ions = number of OH- ions.
• [H+] = [OH-] = 1
10,000,000 M = 10-7 M
• Note that brackets indicate molar concentration.
Chapter 3 Water and the Fitness of the Environment 29
A solution in which:
• [H+] = [OH-] is a neutral solution.
• [H+] > [OH-] is an acidic solution.
• [H+] < [OH-] is a basic solution.
Strong acids and bases dissociate completely in water.
• Example: HCl and NaOH
• Single arrows indicate complete dissociation.
NaOH Na+ + OH
Weak acids and bases dissociate only partially and reversibly.
• Examples: NH3 (ammonia) and H2CO3 (carbonic acid)
• Double arrows indicate a reversible reaction; at equilibrium there will be a fixed ratio of reactants and products.
H2CO3 HCO3- H+
Carbonic Bicarbonate + Hydrogen
acid ion ion
2. The pH scale
In any aqueous solution:
[H+][OH-] = 1.0 ⋅ 10-14
For example:
• In a neutral solution, [H+] = 10-7 M and [OH-] = 10-7 M.
• In an acidic solution where the [H+] = 10-5 M, the [OH-] = 10-9 M. • In a basic solution where the [H+] = 10-9 M, the [OH-] = 10-5 M. pH scale = Scale used to measure degree of acidity. It ranges from 0 to 14. pH = Negative log10 of the [H+] expressed in moles per liter.
• pH of 7 is a neutral solution.
• pH < 7 is an acidic solution.
• pH > 7 is a basic solution.
30 Unit I The Chemistry of Life
• Most biological fluids are within the pH range of 6 to 8. There are some exceptions such as stomach acid with pH = 1.5. (See Campbell, Figure 3.9) • Each pH unit represents a tenfold difference (scale is logarithmic), so a slight change in pH represents a large change in actual [H+].
3. Buffers
By minimizing wide fluctuations in pH, buffers help organisms maintain the pH of body fluids within the narrow range necessary for life (usually pH 6-8).
Buffer = Substance that minimizes large sudden changes in pH.
• Are combinations of H+-donor and H+-acceptor forms in a solution of weak acids or bases
• Work by accepting H+ ions from solution when they are in excess and by donating H+ ions to the solution when they have been depleted
Example: Bicarbonate buffer
response to a
rise in pH
H2CO3 HCO3-+ H+
H+ donor response to a H+ acceptor Hydrogen
(weak acid) drop in pH (weak base) ion
HCl + NaHCO3 H2CO3 + NaCl
strong weak
acid acid
NaOH + H2CO3 NaHCO3 + H2O
strong weak
base base
III. Acid Precipitation Threatens the Fitness of the Environment
Acid precipitation = Rain, snow, or fog more strongly acidic than pH 5.6.
• Has been recorded as low as pH 1.5 in West Virginia
• Occurs when sulfur oxides and nitrogen oxides in the atmosphere react with water in the air to form acids which fall to Earth in precipitation
• Major oxide source is the combustion of fossil fuels by industry and cars • Acid rain affects the fitness of the environment to support life.
• Lowers soil pH which affects mineral solubility. May leach out necessary mineral nutrients and increase the concentration of minerals that are potentially toxic to vegetation in higher concentration (e.g., aluminum). This is contributing to the decline of some European and North American forests.
Chapter 3 Water and the Fitness of the Environment 31
• Lowers the pH of lakes and ponds, and runoff carries leached out soil minerals into aquatic ecosystems. This adversely affects aquatic life. Example: In the Western Adirondack Mountains, there are lakes with a pH < 5 that have no fish.
What can be done to reduce the problem?
• Add industrial pollution controls.
• Develop and use antipollution devices.
• Increase involvement of voters, consumers, politicians, and business leaders.
REFERENCES
Campbell, N., et al. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Gould, R. Going Sour: Science and Politics of Acid Rain. Boston: Birkhauser, 1985. Henderson, L. J. The Fitness of the Environment. Boston: Beacon Press, 1958. Mohnen, V.A. "The Challenge of Acid Rain." Scientific American, August 1988.
CHAPTER 4
CARBON AND
MOLECULAR DIVERSITY
OUTLINE
I. The Importance of Carbon
A. Organic chemistry is the study of carbon compounds
B. Carbon atoms are the most versatile building blocks of molecules
C. Variation in carbon skeletons contributes to the diversity of organic molecules II. Functional Groups
A. Functional groups also contribute to the molecular diversity of life
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Summarize the philosophies of vitalism and mechanism, and explain how they influenced the development of organic chemistry, as well as mainstream biological thought.
2. Explain how carbon’s electron configuration determines the kinds and number of bonds carbon will form.
3. Describe how carbon skeletons may vary, and explain how this variation contributes to the diversity and complexity of organic molecules.
4. Distinguish among the three types of isomers: structural, geometric and enantiomers. 5. Recognize the major functional groups, and describe the chemical properties of organic molecules in which they occur.
KEY TERMS
organic chemistry enantiomer aldehyde amine hydrocarbon functional group ketone sulfhydryl group isomer hydroxyl group carboxyl group thiol structural isomer alcohol carboxylic acid phosphate group geometric isomer carbonyl group amino group
LECTURE NOTES
Aside from water, most biologically important molecules are carbon-based (organic). The structural and functional diversity of organic molecules emerges from the ability of carbon to form large, complex and diverse molecules by bonding to itself and to other elements such as H, O, N, S, and P.
34 Unit I The Chemistry of Life
I. The Importance of Carbon
A. Organic chemistry is the study of carbon compounds
Organic chemistry = The branch of chemistry that specializes in the study of carbon compounds.
Organic molecules = Molecules that contain carbon
Vitalism = Belief in a life force outside the jurisdiction of chemical/physical laws. • Early 19th century organic chemistry was built on a foundation of vitalism because organic chemists could not artificially synthesize organic compounds. It was believed that only living organisms could produce organic compounds. Mechanism = Belief that all natural phenomena are governed by physical and chemical laws.
• Pioneers of organic chemistry began to synthesize organic compounds from inorganic molecules. This helped shift mainstream biological thought from vitalism to mechanism.
• For example, Friedrich Wohler synthesized urea in 1828; Hermann Kolbe synthesized acetic acid.
• Stanley Miller (1953) demonstrated the possibility that organic compounds could have been produced under the chemical conditions of primordial Earth.
B. Carbon atoms are the most versatile building blocks of molecules The carbon atom:
• Usually has an atomic number of 6; therefore, it has 4 valence electrons. • Usually completes its outer energy shell by sharing valence electrons in four covalent bonds. (Not likely to form ionic bonds.)
Emergent properties, such as the kinds and number of bonds carbon will form, are determined by their tetravalent electron configuration.
• It makes large, complex molecules possible. The carbon atom is a central point from which the molecule branches off into four directions.
• It gives carbon covalent compatibility with many different elements. The four major atomic components of organic molecules are as follows:
• It determines an organic molecule’s three-dimensional shape, which may affect molecular function. For example, when carbon forms four single covalent bonds, the four valence orbitals hybridize into teardrop-shaped orbitals that angle from the carbon atoms toward the corners of an imaginary tetrahedron.
Chapter 4 Carbon and Molecular Diversity 35
C. Variation in carbon skeletons contributes to the diversity of organic molecules
Covalent bonds link carbon atoms together in long chains that form the skeletal framework for organic molecules. These carbon skeletons may vary in: • Length
• Shape (straight chain, branched, ring)
• Number and location of double bonds
• Other elements covalently bonded to available sites
This variation in carbon skeletons contributes to the complexity and diversity of organic molecules (see Campbell, Figure 4.4).
Hydrocarbons = Molecules containing only carbon and hydrogen
• Are major components of fossil fuels produced from the organic remains of organisms living millions of years ago, though they are not prevalent in living organisms.
• Have a diversity of carbon skeletons which produce molecules of various lengths and shapes.
• As in hydrocarbons, a carbon skeleton is the framework for the large diverse organic molecules found in living organisms. Also, some biologically important molecules may have regions consisting of hydrocarbon chains (e.g. fats).
• Hydrocarbon chains are hydrophobic because the C−C and C−H bonds are nonpolar.
1. Isomers
Isomers = Compounds with the same molecular formula but with different structures and hence different properties. Isomers are a source of variation among organic molecules.
There are three types of isomers (see Campbell, Figure 4.6):
Structural isomers = Isomers that differ in the covalent arrangement of their atoms.
H|
H−C−H
H H H H H H
| | | | | |
H−C−C−C−C−H H−C−C−C−H
| | | | | | |
H H H H H H H
• Number of possible isomers increases as the carbon skeleton size increases.
• May also differ in the location of double bonds.
Geometric isomers = Isomers which share the same covalent partnerships, but differ in their spatial arrangements.
HO OH H OH
\ / \ /
C = C C = C
/ \ / \
H H HO H
• Result from the fact that double bonds will not allow the atoms they join to rotate freely about the axis of the bonds.
• Subtle differences between isomers affects their biological activity.
36 Unit I The Chemistry of Life
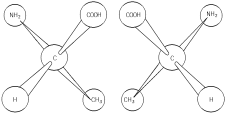
Enantiomers = Isomers that are mirror images of each other.
• Can occur when four different atoms or groups of atoms are bonded to the same carbon (asymmetric carbon).
• There are two different spatial arrangements of the four groups around the asymmetric carbon. These arrangements are mirror images.
• Usually one form is biologically active and its mirror image is not.
II. Functional Groups
A. Functional groups also contribute to the molecular diversity of life Small characteristic groups of atoms (functional groups) are frequently bonded to the carbon skeleton of organic molecules. These functional groups:
• Have specific chemical and physical properties.
• Are the regions of organic molecules which are commonly chemically reactive. • Behave consistently from one organic molecule to another.
• Depending upon their number and arrangement, determine unique chemical properties of organic molecules in which they occur.
As with hydrocarbons, diverse organic molecules found in living organisms have carbon skeletons. In fact, these molecules can be viewed as hydrocarbon derivatives with functional groups in place of H, bonded to carbon at various sites along the molecule. 1. The hydroxyl group
Hydroxyl group = A functional group that consists of a hydrogen atom bonded to an oxygen atom, which in turn is bonded to carbon (−OH).
• Is a polar group; the bond between the oxygen and hydrogen is a polar covalent bond.
• Makes the molecule to which it is attached water soluble. Polar water molecules are attracted to the polar hydroxyl group which can form hydrogen bonds.
• Organic compounds with hydroxyl groups are called alcohols.
2. The carbonyl group
Carbonyl group = Functional group that consists of a carbon atom double-bonded to oxygen (−CO).
• Is a polar group. The oxygen can be involved in hydrogen bonding, and molecules with his functional group are water soluble.
• Is a functional group found in sugars.
Chapter 4 Carbon and Molecular Diversity 37
• If the carbonyl is at the end off the carbon skeleton, the compound is an aldehyde.
OH OH O
| | //
H−C C C
| | |
H H H
Glyceraldehyde
• If the carbonyl is at the end of the carbon skeleton, the compound is a ketone.
H O H
| |
H−C C C−H
| |
H H
Acetone
3. The carboxyl group
Carboxyl group = Functional group that consists of a carbon atom which is both double-bonded to an oxygen and single-bonded to the oxygen of a hydroxyl group (−COOH).
• Is a polar group and water soluble. The covalent bond between oxygen and hydrogen is so polar, that the hydrogen reversibly dissociates as H+. This polarity results from the combined effect of the two electronegative oxygen atoms bonded to the same carbon.
H O H O
| // | //
H−C−C H−C−C + H+
| \ | \
H OH H O
Acetic Acetate Hydrogen
acid ion ion
• Since it donates protons, this group has acidic properties. Compounds with this functional group are called carboxylic acids.
4. The amino group
Amino group = Functional group that consists of a nitrogen atom bonded to two hydrogens and to the carbon skeleton (−NH2).
• Is a polar group and soluble in water.
• Acts as a weak base. The unshared pair of electrons on the nitrogen can accept a proton, giving the amino group a +1 charge.
H H
/ /
R−N + H+ R−+N−H
\ \
H H
Amine Ammonium
ion
• Organic compounds with this function group are called amines. 5. The Sulfhydryl group
Sulfhydryl group = Functional group which consists of an atom of sulfur bonded to an atom of hydrogen (−SH).
38 Unit I The Chemistry of Life
• Help stabilize the structure of proteins. (Disulfide bridges will be discussed with tertiary structure of proteins in Chapter 5, Structure and Function of Macromolecules.)
• Organic compounds with this functional group are called thiols.
6. The phosphate group
Phosphate group = Functional group which is the dissociated form of phosphoric acid (H3PO4).
• Loss of two protons by dissociation leaves the phosphate group with a negative charge.
O O
R−O−P−OH R−O−P−O-+ 2H+
| |
OH O-
• Has acid properties since it loses protons.
• Polar group and soluble in water.
• Organic phosphates are important in cellular energy storage and transfer. (ATP is discussed with energy for cellular work in Chapter 6: Introduction to Metabolism.)
REFERENCES
Campbell, N. et al. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Lehninger, A.L., D.L. Nelson and M.M. Cox. Principles of Biochemistry. 2nd ed. New York: Worth, 1993.
Whitten, K.W. and K.D. Gailey. General Chemistry. 4th ed. New York: Saunders, 1992.
CHAPTER 5
THE STRUCTURE AND FUNCTION
OF MACROMOLECULES
OUTLINE
I. Polymer Principles
A. Most macromolecules are polymers
B. A limitless variety of polymers can be built from a small set of monomers II. Carbohydrates: Fuel and Building Material
A. Sugars, the smallest carbohydrates, serve as fuel and carbon sources
B. Polysaccharides, the polymers of sugars, have storage and structural roles III. Lipids: Diverse Hydrophobic Molecules
A. Fats store large amounts of energy
B. Phospholipids are major components of cell membranes
C. Steroids include cholesterol and certain hormones
IV. Proteins: The Molecular Tools of the Cell
A. A polypeptide is a polymer of amino acids connected in a specific sequence B. A protein’s function depends on its specific conformation
V. Nucleic Acids: Informational Polymers
A. Nucleic acids store and transmit hereditary information
B. A nucleic acid strand is a polymer of nucleotides
C. Inheritance is based on replication of the DNA double helix
D. We can use DNA and proteins as tape measures of evolution
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. List the four major classes of biomolecules.
2. Explain how organic polymers contribute to biological diversity.
3. Describe how covalent linkages are formed and broken in organic polymers. 4. Describe the distinguishing characteristics of carbohydrates, and explain how they are classified.
5. List four characteristics of a sugar.
6. Identify a glycosidic linkage and describe how it is formed.
7. Describe the important biological functions of polysaccharides.
8. Distinguish between the glycosidic linkages found in starch and cellulose, and explain why the difference is biologically important.
9. Explain what distinguishes lipids from other major classes of macromolecules. 10. Describe the unique properties, building block molecules and biological importance of the three important groups of lipids: fats, phospholipids and steroids.
11. Identify an ester linkage and describe how it is formed.
40 Unit I The Chemistry of Life
12. Distinguish between a saturated and unsaturated fat, and list some unique emergent properties that are a consequence of these structural differences.
13. Describe the characteristics that distinguish proteins from the other major classes of macromolecules, and explain the biologically important functions of this group. 14. List and recognize four major components of an amino acid, and explain how amino acids may be grouped according to the physical and chemical properties of the side chains.
15. Identify a peptide bond and explain how it is formed.
16. Explain what determines protein conformation and why it is important. 17. Define primary structure and describe how it may be deduced in the laboratory. 18. Describe the two types of secondary protein structure, and explain the role of hydrogen bonds in maintaining the structure.
19. Explain how weak interactions and disulfide bridges contribute to tertiary protein structure.
20. Using collagen and hemoglobin as examples, describe quaternary protein structure. 21. Define denaturation and explain how proteins may be denatured.
22. Describe the characteristics that distinguish nucleic acids from the other major groups of macromolecules.
23. Summarize the functions of nucleic acids.
24. List the major components of a nucleotide, and describe how these monomers are linked together to form a nucleic acid.
25. Distinguish between a pyrimidine and a purine.
26. List the functions of nucleotides.
27. Briefly describe the three-dimensional structure of DNA.
KEY TERMS
polymer cellulose polypeptide quaternary structure monomer chitin amino acid denaturation condensation reaction lipid protein chaperone proteins dehydration reaction fat conformation gene hydrolysis fatty acid peptide bond nucleic acid carbohydrate triacylglycerol primary structure deoxyribonucleic acid monosaccharide saturated fatty acid secondary structure ribonucleic acid disaccharide unsaturated fatty acid alpha ( ) helix nucleotide glycosidic linkage steroid pleated sheet pyrimidine polysaccharide cholesterol tertiary structure purine starch protein hydrophobic interaction ribose glycogen conformation disulfide bridges polynucleotide double helix
LECTURE NOTES
The topic of macromolecules lends itself well to illustrate three integral themes that permeate the text and course:
1. There is a natural hierarchy of structural level in biological organization. 2. As one moves up the hierarchy, new properties emerge because of interactions among subunits at the lower levels.
3. Form fits function.
Chapter 5 The Structure and Function of Macromolecules 41
I. Polymer Principles
A. Most macromolecules are polymers
Polymer = (Poly = many; mer = part); large molecule consisting of many identical or similar subunits connected together.
Monomer = Subunit or building block molecule of a polymer
Macromolecule = (Macro = large); large organic polymer
• Formation of macromolecules from smaller building block molecules represents another level in the hierarchy of biological organization.
• There are four classes of macromolecules in living organisms:
1. Carbohydrates
2. Lipids
3. Proteins
4. Nucleic acids
Most polymerization reactions in living organisms are condensation reactions. • Polymerization reactions = Chemical reactions that link two or more small molecules to form larger molecules with repeating structural units.
• Condensation reactions = Polymerization reactions during which monomers are covalently linked, producing net removal of a water molecule for each covalent linkage.
• One monomer loses a hydroxyl (–OH), and the other monomer loses a hydrogen (–H).
• Removal of water is actually indirect, involving the formation of “activated” monomers (discussed in Chapter 6, Introduction t o Metabolism).
• Process requires energy.
• Process requires biological catalysts or enzymes.
Hydrolysis = (Hydro = water; lysis = break); a reaction process that breaks covalent bonds between monomers by the addition of water molecules.
• A hydrogen from the water bonds to one monomer, and the hydroxyl bonds to the adjacent monomer.
• Example: Digestive enzymes catalyze hydrolytic reactions which break apart large food molecules into monomers that can be absorbed into the bloodstream.
B. An immense variety of polymers can be built from a small set of monomers Structural variation of macromolecules is the basis for the enormous diversity of life. • There is unity in life as there are only about 40 to 50 common monomers used to construct macromolecules.
• There is diversity in life as new properties emerge when these universal monomers are arranged in different ways.
II. Carbohydrates: Fuel and Building Material
A. Sugars, the smallest carbohydrates, serve as fuel and carbon sources Carbohydrates = Organic molecules made of sugars and their polymers
• Monomers or building block molecules are simple sugars called monosaccharides.
• Polymers are formed by condensation reactions.
• Carbohydrates are classified by the number of simple sugars.
42 Unit I The Chemistry of Life
1. Monosaccharides
Monosaccharides = (Mono = single; sacchar = sugar); simple sugar in which C, H, and O occur in the ratio of (CH2O).
• Are major nutrients for cells; glucose is the most common
• Can be produced (glucose) by photosynthetic organisms from CO2, H2O, and sunlight
• Store energy in their chemical bonds which is harvested by cellular respiration
• Their carbon skeletons are raw material for other organic molecules.
• Can be incorporated as monomers into disaccharides and polysaccharides Characteristics of a sugar:
a. An –OH group is attached to each carbon except one, which is double bonded to an oxygen (carbonyl).
Aldehyde Ketone
Terminal carbon forms a Carbonyl group is within
double bond with oxygen. the carbon skeleton.
H O H
\ // |
C H−C−OH
| |
H−C−OH C=O
| |
HO−C−H HO−C−H
| |
H−C−OH H−C−OH
| |
H−C−OH H−C−OH
| |
H−C−OH H−C−OH
| |
H H
Glucose Fructose
(aldose) (ketose)
b. Size of the carbon skeleton varies from three to seven carbons. The most common monosaccharides are:
Chapter 5 The Structure and Function of Macromolecules 43
c. Spatial arrangement around asymmetric carbons may vary. For example, glucose and galactose are enantiomers.
H O H O
\ / / \ / /
C C
H−C−OH H−C−OH
HO−C−H HO− C−H
H C OH HO C H
H−C−OH H−C−OH
H−C−OH H−C−OH
H H
Glucose Galactose
The small difference between isomers affects molecular shape which gives these molecules distinctive biochemical properties.
d. In aqueous solutions, many monosaccharides form rings. Chemical equilibrium favors the ring structure.
H O
\ //
C
H C OH
HO C H
H C OH
H C OH
H C OH
H
Linear Form
of Glucose
2. Disaccharides
Ring Form of Glucose
Disaccharide = (Di = two; sacchar = sugar); a double sugar that consists of two monosaccharides joined by a glycosidic linkage.
Glycosidic linkage = Covalent bond formed by a condensation reaction between two sugar monomers; for example, maltose:
44 Unit I The Chemistry of Life

Examples of disaccharides include:
B. Polysaccharides, the polymers of sugars, have storage and structural roles Polysaccharides = Macromolecules that are polymers of a few hundred or thousand monosaccharides.
• Are formed by linking monomers in enzyme-mediated condensation reactions • Have two important biological functions:
1. Energy storage (starch and glycogen)
2. Structural support (cellulose and chitin)
1. Storage polysaccharides
Cells hydrolyze storage polysaccharides into sugars as needed. Two most common storage polysaccharides are starch and glycogen.
Starch = Glucose polymer that is a storage polysaccharide in plants.
• Helical glucose polymer with 1-4 linkages (see Campbell, Figure 5.6) • Stored as granules within plant organelles called plastids
• Amylose, the simplest form, is an unbranched polymer.
• Amylopectin is branched polymer.
• Most animals have digestive enzymes to hydrolyze starch.
• Major sources in the human diet are potato tubers and grains (e.g., wheat, corn, rice, and fruits of other grasses).
Chapter 5 The Structure and Function of Macromolecules 45
Glycogen = Glucose polymer that is a storage polysaccharide in animals. • Large glucose polymer that is more highly branched ( 1-4 and 4-6 linkages) than amylopectin
• Stored in the muscle and liver of humans and other vertebrates 2. Structural polysaccharides
Structural polysaccharides include cellulose and chitin.
Cellulose = Linear unbranched polymer of D-glucose in ( 1-4, 4-6) linkages. • A major structural component of plant cell walls
• Differs from starch (also a glucose polymer) in its glycosidic linkages (see Campbell, Figure 5.7)

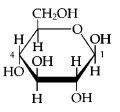
• Cellulose and starch have different three-dimensional shapes and properties as a result of differences in glycosidic linkages.
• Cellulose reinforces plant cell walls. Hydrogen bonds hold together parallel cellulose molecules in bundles of microfibrils (see Campbell, Figure 5.8) • Cellulose cannot be digested by most organisms, including humans, because they lack an enzyme that can hydrolyze the 1-4 linkage. (Exceptions are some symbiotic bacteria, other microorganisms and some fungi.)
Chitin = A structural polysaccharide that is a polymer of an amino sugar (see Campbell, Figure 5.9).
• Forms exoskeletons of arthropods

• Found as a building material in the cell walls of some
fungi
• Monomer is an amino sugar, which is similar t o
beta-glucose with a nitrogen-containing group
replacing the hydroxyl on carbon 2.
46 Unit I The Chemistry of Life
III. Lipids: Diverse Hydrophobic Molecules
Lipids = Diverse group of organic compounds that are insoluble in water, but will dissolve in nonpolar solvents (e.g., ether, chloroform, benzene). Important groups are fats, phospholipids, and steroids.
A. Fats store large amounts of energy
Fats = Macromolecules are constructed from (see Campbell, Figure 5.10): 1. Glycerol, a three-carbon alcohol
2. Fatty acid (carboxylic acid)
• Composed of a carboxyl group at one end and an attached hydrocarbon chain (“tail”)
• Carboxyl functional group (“head”) has properties of an acid.
• Hydrocarbon chain has a long carbon skeleton usually with an even number of carbon atoms (most have 16 – 18 carbons).
• Nonpolar C–H bonds make the chain hydrophobic and not water soluble.
O H H H H H H H
During the formation of a fat, enzyme- \\
catalyzed condensation reactions link H C – C – C– C– C – C – C– C–H glycerol to fatty acids by an ester /
linkage. H−C−OH HO H H H H H H H Fatty acid
H−C−OH
Ester linkage = Bond formed between a
hydroxyl group and a carboxyl group. H−C−OH ÂH2O
H−C−OH
H
Glycerol
Each of glycerol’s three hydroxyl groups Ester linkage
can bond to a fatty acid by an ester H O H H H H H H H linkage producing a fat.
H C O− C− C C C C− C C C H
Triacylglycerol = A fat composed of H C−OH H H H H H H H three fatty acids bonded to one glycerol
by ester linkages (triglyceride). H C−OH
H
Some characteristics of fat include:
• Fats are insoluble in water. The long fatty acid chains are hydrophobic because of the many nonpolar C–H bonds.
• The source of variation among fat molecules is the fatty acid composition. • Fatty acids in a fat may all be the same, or some (or all) may differ.
• Fatty acids may vary in length.
• Fatty acids may vary in the number and location of carbon-to-carbon double bonds.
Chapter 5 The Structure and Function of Macromolecules 47
• In many commercially prepared food products, unsaturated fats are artificially hydrogenated to prevent them from separating out as oil (e.g., peanut butter and margarine).
Fat serves many useful functions:
• Energy storage. One gram of fat stores twice as much energy as a gram of polysaccharide. (Fat has a higher proportion of energy rich C–H bonds.) • More compact fuel reservoir than carbohydrate. Animals store more energy with less weight than plants which use starch, a bulky form of energy storage. • Cushions vital organs in mammals (e.g., kidney).
• Insulates against heat loss (e.g., in mammals such as whales and seals). B. Phospholipids
Phospholipids = Compounds with molecular building blocks of glycerol, two fatty acids, a phosphate group, and usually, an additional small chemical group attached to the phosphate (see Campbell , Figure 5.12)
• Differ from fat in that the third carbon of glycerol is joined to a negatively charged phosphate group
• Can have small variable molecules (usually charged or polar) attached t o phosphate
• Are diverse depending upon differences in fatty acids and in phosphate attachments
• Show ambivalent behavior toward water. Hydrocarbon tails are hydrophobic and the polar head (phosphate group with attachments) is hydrophilic.
• Cluster in water as their hydrophobic portions turn away from water. One such cluster, a micelle, assembles so the hydrophobic tails turn toward the water-free interior and the hydrophilic phosphate heads arrange facing outward in contact with water (see Campbell, Figure 5.13).
• Are major constituents of cell membranes. At the cell surface, phospholipids form a bilayer held together by hydrophobic interactions among the hydrocarbon tails. Phospholipids in water will spontaneously form such a bilayer.
C. Steroids
Steroids = Lipids which have four fused carbon rings with various functional groups attached.
48 Unit I The Chemistry of Life
Cholesterol is an important steroid.
• Is the precursor to many other steroids including
vertebrate sex hormones and bile acids.
• Is a common component of animal cell membranes.
• Can contribute to atherosclerosis.
IV. Proteins: The Molecular Tools of the Cell
Polypeptide chains = Polymers of amino acids that are
arranged in a specific linear sequence and are linked by
peptide bonds.
Protein = A macromolecule that consists of one or more polypeptide chains folded and coiled into specific conformations.
• Are abundant, making up 50% or more of cellular dry weight
• Have important and varied functions in the cell:
1. Structural support
2. Storage (of amino acids)
3. Transport (e.g., hemoglobin)
4. Signaling (chemical messengers)
5. Cellular response to chemical stimuli (receptor proteins)
6. Movement (contractile proteins)
7. Defense against foreign substances and disease-causing organisms (antibodies)
8. Catalysis of biochemical reactions (enzymes)
• Vary extensively in structure; each type has a unique three-dimensional shape (conformation)
• Though they vary in structure and function, they are commonly made of only 20 amino acid monomers.
A. A polypeptide is a polymer of amino acids connected in a specific sequence Amino acid = Building block molecule of a protein; most consist of an asymmetric carbon, termed the alpha carbon, which is covalently bonded to a(n):
1. Hydrogen atom.
2. Carboxyl group.
3. Amino group.
4. Variable R group (side chain) specific to each amino acid. Physical and chemical properties of the side chain determine the uniqueness of each amino acid.
H O
| //
H3N+−C−C
| \
R O-
(At pH’s normally found in the cell, both the carboxyl and amino groups are ionized.)
Amino acids contain both carboxyl and amino functional groups. Since one group acts as a weak acid and the other group acts as a weak base, an amino acid can exist in three ionic states. The pH of the solution determines which ionic state predominates.
H H O H H O H H O | | // as pH increases | | // as pH increases \ | // H−N+−C−C H−N+−C−C + H+ N−C−C + H+ | | \ as pH decreases | | \ as pH decreases / | \ H H OH H H O- H H O
Cation Zwitterion Anion (dipolar ion)
Chapter 5 The Structure and Function of Macromolecules 49
The twenty common amino acids can be grouped by properties of side chains (see Campbell, Figure 5.15):
1. Nonpolar side groups (hydrophobic). Amino acids with nonpolar groups are less soluble in water.
2. Polar side groups (hydrophilic). Amino acids with polar side groups are soluble in water. Polar amino acids can be grouped further into:
a. Uncharged polar
b. Charged polar
• Acidic side groups. Dissociated carboxyl group gives these side groups a negative charge.
• Basic side groups. An amino group with an extra proton gives these side groups a net positive charge.
Polypeptide chains are polymers that are formed when amino acid monomers are linked by peptide bonds (see Campbell, Figure 5.16).
R R
Peptide bond = Covalent bond formed by a | | condensation reaction that links the carboxyl H−N−C−C−OH H−N−C−C−OH group of one amino acid to the amino group | | | | | | | | of another. H H O H H O
• Has polarity with an amino group on
amino acid amino acid
one end (N-terminus) and a carboxyl group  H2O on the other (C-terminus). R R
| |
−C−C N−C• Has a backbone of the repeating | || |sequence −N−C−C−N−C−C−. H O
N-terminus C-terminus
Peptide bond
Polypeptide chains:
• Range in length from a few monomers to more than a thousand.
• Have unique linear sequences of amino acids.
B. A protein’s function depends on its specific conformation
A protein’s function depends upon its unique conformation.
Protein conformation = Three-dimensional shape of a protein.
Native conformation = Functional conformation of a protein found under normal biological conditions.
• Enables a protein to recognize and bind specifically to another molecule (e.g., hormone/receptor, enzyme/substrate, and antibody/antigen)
• Is a consequence of the specific linear sequence of amino acids in the polypeptide
• Is produced when a newly formed polypeptide chain coils and folds spontaneously, mostly in response to hydrophobic interactions
• Is stabilized by chemical bonds and weak interactions between neighboring regions of the folded protein
1. Four levels of protein structure
The correlation between form and function in proteins is an emergent property resulting from superimposed levels of protein structure (see Campbell, Figure 5.24): • Primary structure
• Secondary structure
50 Unit I The Chemistry of Life
• Tertiary structure
• When a protein has two or more polypeptide chains, it also has quaternary structure.
a. Primary structure
Primary structure = Unique sequence of amino acids in a protein.
• Determined by genes
• Slight change can affect a protein’s conformation and function (e.g., sickle-cell hemoglobin; see Campbell, Figure 5.19).
• Can be sequenced in the laboratory. A pioneer in this work was Frederick Sanger who determined the amino acid sequence in insulin
(late 1940s and early 1950s). This laborious process involved:
1) Determination of amino acid composition by complete acid
hydrolysis of peptide bonds and separation of resulting amino acids
by chromatography. Using these techniques, Sanger identified the
amino acids and determined the relative proportions of each.
2) Determination of amino acid sequence by partial hydrolysis with enzymes and other catalysts to break only specific peptide bonds.
Sanger deductively reconstructed the primary structure from fragments
with overlapping segments.
• Most of the sequencing process is now automated.
b. Secondary structure
Secondary structure = Regular, repeated coiling and folding of a protein’s
polypeptide backbone (see Campbell, Figure 5.20).
• Contributes to a protein’s overall conformation.
• Stabilized by hydrogen bonds between peptide linkages in the protein’s backbone (carbonyl and amino groups).
• The major types of secondary structure are alpha ( ) helix and beta ( ) pleated sheet.
1) Alpha ( ) helix
Alpha ( ) helix = Secondary structure of a polypeptide that is a helical
coil stabilized by hydrogen bonding between every fourth peptide bond
(3.6 amino acids per turn).
• Described by Linus Pauling and Robert Corey in 1951.
• Found in fibrous proteins (e.g., -keratin and collagen) for most
of their length and in some portions of globular proteins.
2) Beta ( ) pleated sheet
Beta ( ) pleated sheet = Secondary protein structure which is a sheet of
antiparallel chains folded into accordion pleats.
• Parallel regions are held together by either intrachain or
interchain hydrogen bonds (between adjacent polypeptides).
• Make up the dense core of many globular proteins (e.g.,
lysozyme) and the major portion of some fibrous proteins (e.g.,
fibroin, the structural protein of silk).
c. Tertiary structure
Tertiary structure = The three-dimensional shape of a protein. The irregular contortions of a protein are due to bonding between and among side chains (R groups) and to interaction between R groups and the aqueous environment (see Campbell, Figure 5.22).
Types of bonds contributing to tertiary structure are weak interactions and covalent linkage (both may occur in the same protein).
Chapter 5 The Structure and Function of Macromolecules 51
1) Weak interactions
Protein shape is stabilized by the cumulative effect of weak
interactions. These weak interactions include:
• Hydrogen bonding between polar side chains.
• Ionic bonds between charged side chains.
• Hydrophobic interactions between nonpolar side chains in protein’s interior.
Hydrophobic interactions = (Hydro = water; phobos = fear); the
clustering of hydrophobic molecules as a result of their mutual exclusion from water.
2) Covalent linkage
Disulfide bridges form between two cysteine monomers brought together by folding of the protein. This is a strong bond that reinforces
conformation.
H O
| // | |
H3N+−C−C −C−CH2−S S−CH2−C−
| \ | |
CH2 O-
|
SH
Cysteine Disulfide Bridge
(S of one cysteine sulfhydryl, bonds
to the S of a second cysteine.)
d. Quaternary structure
Quaternary structure = Structure that results from the interactions between and among several polypeptides chains (subunits) (see Campbell, Figure 5.23). • Example: Collagen, a fibrous protein with three helical polypeptides supercoiled into a triple helix; found in animal connective tissue, collagen’s supercoiled quaternary structure gives it strength.
• Some globular proteins have subunits that fit tightly together. Example: Hemoglobin, a globular protein that has four subunits (two chains and two chains)
2. What determines protein conformation?
A protein’s three-dimensional shape is a consequence of the interactions responsible for secondary and tertiary structure.
• This conformation is influenced by physical and chemical environmental conditions.
• If a protein’s environment is altered, it may become denatured and lose its native conformation.
Denaturation = A process that alters a protein’s native conformation and biological activity. Proteins can be denatured by:
• Transfer to an organic solvent. Hydrophobic side chains, normally inside the protein’s core, move towards the outside. Hydrophilic side chains turn away from the solvent towards the molecule’s interior.
• Chemical agents that disrupt hydrogen bonds, ionic bonds and disulfide bridges.
• Excessive heat. Increased thermal agitation disrupts weak interactions (see Campbell, Figure 5.25).
52 Unit I The Chemistry of Life
The fact that some denatured proteins return to their native conformation when environmental conditions return to normal is evidence that a protein’s amino acid sequence (primary structure) determines conformation. It influences where and which interactions will occur as the molecule arranges into secondary and tertiary structure.
3. The protein-folding problem
Even though primary structure ultimately determines a protein’s conformation, three-dimensional shape is difficult to predict solely on the basis of amino acid sequence. It is difficult to find the rules of protein folding because:
• Most proteins pass through several intermediate stages in the folding process; knowledge of the final conformation does not reveal the folding process required to create it.
• A protein’s native conformation may be dynamic, alternating between several shapes.
Using recently developed techniques, researchers hope to gain new insights into protein folding:
• Biochemists can now track a protein as it passes through its intermediate stages during the folding process.
• Chaperone proteins have just been discovered that temporarily brace a folding protein.
Rules of protein folding are important to molecular biologists and the
biotechnology industry. This knowledge should allow the design of proteins for specific purposes.
V. Nucleic Acids: Informational Polymers
A. Nucleic acids store and transmit hereditary information
Protein conformation is determined by primary structure. Primary structure, in turn, is determined by genes; hereditary units that consist of DNA, a type of nucleic acid. There are two types of nucleic acids.
1. Deoxyribonucleic acid (DNA)
• Contains coded information that programs all cell activity.
• Contains directions for its own replication.
• Is copied and passed from one generation of cells to another.
• In eukaryotic cells, is found primarily in the nucleus.
• Makes up genes that contain instructions for protein synthesis. Genes do not directly make proteins, but direct the synthesis of mRNA.
2. Ribonucleic acid (RNA)
• Functions in the actual synthesis of proteins coded for by DNA.
• Sites of protein synthesis are on ribosomes in the cytoplasm.
• Messenger RNA (mRNA) carries encoded genetic message from the nucleus to the cytoplasm.
• The flow of genetic information goes from DNA RNA protein (see Campbell, Figure 5.26).
Nucleus mRNA Cytoplasm
Genetic message is Genetic message
transcribed from moves into translated into
DNA onto mRNA cytoplasm a protein.
Chapter 5 The Structure and Function of Macromolecules 53
B. A nucleic acid strand is a polymer of nucleotides
Nucleic acid = Polymer of nucleotides linked together by condensation reactions. Nucleotide = Building block molecule of a nucleic acid; made of (1) a five-carbon sugar covalently bonded to (2) a phosphate group and (3) a nitrogenous base. 1. Pentose (5-carbon sugar)
There are two pentoses found in nucleic acids: ribose and deoxyribose.

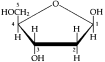
Ribose is the pentose in RNA. Deoxyribose is the pentose in DNA. (It lacks the −OH group at
the number two carbon.)
2. Phosphate
The phosphate group is attached to the number 5 carbon of the sugar. 3. Nitrogenous base
There are two families of nitrogenous bases:
Pyrimidine = Nitrogenous base characterized by a six-membered ring made up of carbon and nitrogen atoms. For example:
• Cytosine (C)
• Thymine (T); found only in DNA
• Uracil (U); found only in RNA
Purine = Nitrogenous base characterized by a five-membered ring fused to a six membered ring. For example:
• Adenine (A)
• Guanine (G)
Nucleotides have various functions:
• Are monomers for nucleic acids.
• Transfer chemical energy from one molecule to another (e.g., ATP). • Are electron acceptors in enzyme-controlled
redox reactions of the cell (e.g., NAD).
A nucleic-acid polymer or polynucleotise, results from joining nucleotides together by covalent bonds called phosphodiester linkages. The bond is formed between the phosphate of one nucleotide and the sugar of the next.
• Results in a backbone with a repeating pattern of sugar-phosphate-sugar-phosphate. • Variable nitrogenous bases are attached to the sugar-phosphate backbone.
• Each gene contains a unique linear sequence of nitrogenous bases which codes for a unique linear sequence of amino acids in a protein.
Phosphate Base
Sugar
54 Unit I The Chemistry of Life
C. Inheritance is based on precise replication of the DNA double helix In 1953, James Watson and Francis Crick proposed the double helix as the three dimensional structure of DNA.
• Consists of two nucleotide chains wound in a double helix.
• Sugar-phosphate backbones are on the outside of the helix.
• The two polynucleotide strands of DNA are held together by hydrogen bonds between the paired nitrogenous bases and by van der Waals attraction between the stacked bases (see Campbell, Figure 5.28).
• Base-pairing rules are that adenine (A) always pairs with thymine (T); guanine (G) always pairs with cytosine (C).
• Two strands of DNA are complimentary and thus can serve as templates to make new complementary strands. It is this mechanism of precise copying that makes inheritance possible.
• Most DNA molecules are long, containing thousands or millions of base pairs. D. We can use DNA and proteins as tape measures of evolution
Closely related species have more similar sequences of DNA and amino acids, than more distantly related species. Using this type of molecular evidence, biologists can deduce evolutionary relationships among species.
REFERENCES
Alberts, B., et al. Essential Cell Biology: An Introduction to the Molecular Biology of the Cell. New York: Garland Publishing, Inc., 1998.
Campbell, N., et al. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Lehninger, A.L., D.L. Nelson and M.M. Cox. Principles of Biochemistry. 2nd ed. New York: Worth, 1993.
Brown, T.L., H. E. Le May, Jr., and B. Bursten. Chemistry: The Central Science. 7th Ed. Upper Saddle River, New Jersey: Prentice Hall, 1997.
CHAPTER 6
AN INTRODUCTION TO
METABOLISM
OUTLINE
I. Metabolism, Energy and Life
A. The chemistry of life is organized into metabolic pathways
B. Organisms transform energy
C. The energy transformations of life are subject to two laws of thermodynamics D. Organisms live at the expense of free energy
E. ATP powers cellular work by coupling exergonic to endergonic reactions II. Enzymes
A. Enzymes speed up metabolic reactions by lowering energy barriers
B. Enzymes are substrate-specific
C. The active site is an enzyme’s catalytic center
D. A cell’s physical and chemical environment affects enzyme activity
III. The Control of Metabolism
A. Metabolic control often demends on allosteric regulation
B. The location of enzymes within a cell helps order metabolism
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Explain the role of catabolic and anabolic pathways in the energy exchanges of cellular metabolism.
2. Distinguish between kinetic and potential energy.
3. Distinguish between open and closed systems.
4. Explain, in their own words, the First and Second Laws of Thermodynamics. 5. Explain why highly ordered living organisms do not violate the Second Law of Thermodynamics.
6. Distinguish between entropy and enthalpy.
7. Write the Gibbs equation for free energy change.
8. Explain how changes in enthalpy, entropy and temperature influence the maximum amount of usable energy that can be harvested from a reaction.
9. Explain the usefulness of free energy.
10. List two major factors capable of driving spontaneous processes.
11. Distinguish between exergonic and endergonic reactions.
12. Describe the relationship between equilibrium and free energy change for a reaction.
56 Unit I The Chemistry of Life
13. Describe the function of ATP in the cell.
14. List the three components of ATP and identify the major class of macromolecules to which it belongs.
15. Explain how ATP performs cellular work.
16. Explain why chemical disequilibrium is essential for life.
17. Describe the energy profile of a chemical reaction including activation energy (EA), free energy change ( G) and transition state.
18. Describe the function of enzymes in biological systems.
19. Explain the relationship between enzyme structure and enzyme specificity. 20. Explain the induced fit model of enzyme function and describe the catalytic cycle of an enzyme.
21. Describe several mechanisms by which enzymes lower activation energy. 22. Explain how substrate concentration affects the rate of an enzyme-controlled reaction. 23. Explain how enzyme activity can be regulated or controlled by environmental conditions, cofactors, enzyme inhibitors and allosteric regulators.
24. Distinguish between allosteric activation and cooperativity.
25. Explain how metabolic pathways are regulated.
KEY TERMS
metabolism first law of thermodynamics catalyst noncompetitive inhibitors catabolic pathways second law of thermodynamics activation energy allosteric site anabolic pathways free energy substrate feedback inhibition bioenergetics exergonic reaction active site cooperativity energy endergonic reaction induced fit entropy kinetic energy energy coupling cofactors spontaneous reaction potential energy ATP coenzymes
thermodynamics phosphorylated intermediate competitive inhibitors
LECTURE NOTES
I. Metabolism, Energy and Life
A. The chemistry of life is organized into metabolic pathways
Metabolism = Totality of an organism's chemical processes (see Campbell, Figure 6.1). • Property emerging from specific molecular interactions within the cell. • Concerned with managing cellular resources: material and energy.
Metabolic reactions are organized into pathways that are orderly series of enzymatically controlled reactions. Metabolic pathways are generally of two types: Catabolic pathways = Metabolic pathways that release energy by breaking down complex molecules to simpler compounds (e.g., cellular respiration which degrades glucose to carbon dioxide and water; provides energy for cellular work).
Anabolic pathways = Metabolic pathways that consume energy to build complicated molecules from simpler ones (e.g., photosynthesis which synthesizes glucose from CO2 and H2O; any synthesis of a macromolecule from its monomers).
Metabolic reactions may be coupled, so that energy released from a catabolic reaction can be used to drive an anabolic one.
Chapter 6 An Introduction to Metabolism 57
B. Organisms transform energy
Energy = Capacity to do work
Kinetic energy = Energy in the process of doing work (energy of motion). For example: • Heat (thermal energy) is kinetic energy expressed in random movement of molecules.
• Light energy from the sun is kinetic energy which powers photosynthesis. Potential energy = Energy that matter possesses because of its location or arrangement (energy of position). For example:
• In the earth's gravitational field, an object on a hill or water behind a dam have potential energy.
• Chemical energy is potential energy stored in molecules because of the arrangement of nuclei and electrons in its atoms.
Energy can be transformed from one form to another. For example:
• Kinetic energy of sunlight can be transformed into the potential energy of chemical bonds during photosynthesis.
• Potential energy in the chemical bonds of gasoline can be transformed into kinetic mechanical energy which pushes the pistons of an engine.
C. The energy transformations of life are subject to two laws of thermodynamics Thermodynamics = Study of energy transformations
First Law of Thermodynamics = Energy can be transferred and transformed, but it cannot be created or destroyed (energy of the universe is constant).
Second Law of Thermodynamics = Every energy transfer or transformation makes the universe more disordered (every process increases the entropy of the universe). Entropy = Quantitative measure of disorder that is proportional to randomness (designated by the letter S).
Closed system = Collection of matter under study which is isolated from its surroundings.
Open system = System in which energy can be transferred between the system and its surroundings.
The entropy of a system may decrease, but the entropy of the system plus its surroundings must always increase. Highly ordered living organisms do not violate the second law because they are open systems. For example, animals:
• Maintain highly ordered structure at the expense of increased entropy of their surroundings.
• Take in complex high energy molecules as food and extract chemical energy to create and maintain order.
• Return to the surroundings simpler low energy molecules (CO2 and water) and heat.
58 Unit I The Chemistry of Life
Energy can be transformed, but part of it is dissipated as heat which is largely unavailable to do work. Heat energy can perform work if there is a heat gradient resulting in heat flow from warmer to cooler.
Combining the first and second laws; the quantity of energy in the universe is constant, but its quality is not.
D. Organisms live at the expense of free energy
1. Free energy: a criterion for spontaneous change
Not all of a system's energy is available to do work. The amount of energy that is available to do work is described by the concept of free energy. Free energy (G) is related to the system's total energy (H) and its entropy (S) in the following way: G = H − TS
where:
G = Gibbs free energy (energy available to do work)
H = enthalpy or total energy
T = temperature in °K
S = entropy
Free energy (G) = Portion of a system's energy available to do work; is the difference between the total energy (enthalpy) and the energy not available for doing work (TS).
The maximum amount of usable energy that can be harvested from a particular reaction is the system's free energy change from the initial to the final state. This change in free energy ( G) is given by the Gibbs-Helmholtz equation at constant temperature and pressure:
G = H − T S
where:
G = change in free energy
H = change in total energy (enthalpy)
S = change in entropy
T = absolute temperature in °K (which is °C + 273)
Significance of free energy:
a. Indicates the maximum amount of a system's energy which is available to do work.
b. Indicates whether a reaction will occur spontaneously or not.
• A spontaneous reaction is one that will occur without additional energy. • In a spontaneous process, G or free energy of a system decreases ( G<0).
Chapter 6 An Introduction to Metabolism 59
• A decrease in enthalpy (– H) and an increase in entropy (+ S) reduce the free energy of a system and contribute to the spontaneity of a process.
• A higher temperature enhances the effect of an entropy change. Greater kinetic energy of molecules tends to disrupt order as the chances for random collisions increase.
• When enthalpy and entropy changes in a system have an opposite effect on free energy, temperature may determine whether the reaction will be spontaneous or not (e.g., protein denaturation by increased temperature).
• High energy systems, including high energy chemical systems, are unstable and tend to change to a more stable state with a lower free energy.
2. Free energy and equilibrium
There is a relationship between chemical equilibrium and the free energy change ( G) of a reaction:
• As a reaction approaches equilibrium, the free energy of the system decreases (spontaneous and exergonic reaction).
• When a reaction is pushed away from equilibrium, the free energy of system increases (non-spontaneous and endergonic reaction).
• When a reaction reaches equilibrium, G = 0, because there is no net change in the system.
3. Free energy and metabolism
a. Reactions can be classified based upon their free energy changes: Exergonic reaction = A reaction that proceeds with a net loss of free energy. Endergonic reaction = An energy-requiring reaction that proceeds with a net gain of free energy; a reaction that absorbs free energy from its surroundings.
If a chemical process is exergonic, the reverse process must be endergonic. For example:
• For each mole of glucose oxidized in the exergonic process of cellular respiration, 2870 kJ are released ( G = −2870 kJ/mol or −686 kcal/mol).
• To produce a mole of glucose, the endergonic process of photosynthesis requires an energy input of 2870 kJ ( G = +2870 kJ/mol or +686 kcal/mol).
60 Unit I The Chemistry of Life
In cellular metabolism, endergonic reactions are driven by coupling them to reactions with a greater negative free energy (exergonic). ATP plays a critical role in this energy coupling.
b. Metabolic disequilibrium
Since many metabolic reactions are reversible, they have the potential to reach equilibrium.
• At equilibrium, G = 0, so the system can do no work.
• Metabolic disequilibrium is a necessity of life; a cell at equilibrium is dead.
• In the cell, these potentially reversible reactions are pulled forward away from equilibrium, because the products of some reactions become
reactants for the next reaction in the metabolic pathway.
• For example, during cellular respiration a steady supply of high energy reactants such as glucose and removal of low energy products such as
CO2 and H2O, maintain the disequilibrium necessary for respiration to
proceed.
E. ATP powers cellular work by coupling exergonic to endergonic reactions ATP is the immediate source of energy that drives most cellular work, which includes: • Mechanical work such as beating of cilia, muscle contraction, cytoplasmic flow, and chromosome movement during mitosis and meiosis.
• Transport work such as pumping substances across membranes.
• Chemical work such as the endergonic process of polymerization.
1. The structure and hydrolysis of ATP
ATP (adenosine triphosphate) = Nucleotide with unstable phosphate bonds that the cell hydrolyzes for energy to drive endergonic reactions. ATP consists of:
• Adenine, a nitrogenous
base.
• Ribose, a five-carbon
sugar.
• Chain of three
phosphate groups.
Chapter 6 An Introduction to Metabolism 61
Unstable bonds between the phosphate groups can be hydrolyzed in an exergonic reaction that releases energy.
• When the terminal phosphate bond is hydrolyzed, a phosphate group is removed producing ADP (adenosine diphosphate).
ATP + H2O ADP + P
• Under standard conditions in the laboratory, this reaction releases −31 kJ/mol (−7.3 kcal/mol).
• In a living cell, this reaction releases −55 kJ/mol (–13 kcal/mol)—about 77% more than under standard conditions.
The terminal phosphate bonds of ATP are unstable, so:
• The products of the hydrolysis reaction are more stable than the reactant. • Hydrolysis of the phosphate bonds is thus exergonic as the system shifts to a more stable state.
2 How ATP performs work
Exergonic hydrolysis of ATP is coupled with endergonic processes by transferring a phosphate group to another molecule.
• Phosphate transfer is enzymatically controlled.
• The molecule acquiring the phosphate (phosphorylated or activated intermediate) becomes more reactive.
For example, conversion of glutamic acid to glutamine (see Campbell, Figure 6.7):
Glu + NH3 Gln G = +14.2 kJ/mol (+3.4 kcal/mol) glutamic acid ammonia glutamine (endergonic)
Two step process of energy coupling with ATP hydrolysis:
1. Hydrolysis of ATP and phosphorylation of glutamic acid.
Glu + ATP Glu– P + ADP
unstable
phosphorylated
intermediate
2. Replacement of the phosphate with the reactant ammonia.
Glu– P + NH3 Gln + P
Overall G:
Glu + NH3 Gln G = + 14.2 kJ/mol
ATP ADP + P G = – 31.0 kJ/mol
Net G=– 16.8 kJ/mol
(Overall process is exergonic)
3. The regeneration of ATP
ATP is continually regenerated by the cell.
• Process is rapid (107 molecules used and regenerated/sec/cell).
• Reaction is endergonic.
ADP + P ATP G = + 31 kJ/mol (+7.3 kcal/mol)
• Energy to drive the endergonic regeneration of ATP comes from the exergonic process of cellular respiration.
II. Enzymes
A. Enzymes speed up metabolic reactions by lowering energy barriers Free energy change indicates whether a reaction will occur spontaneously, but does not give information about the speed of reaction.
62 Unit I The Chemistry of Life
• A chemical reaction will occur spontaneously if it releases free energy (– G), but it may occur too slowly to be effective in living cells.
• Biochemical reactions require enzymes to speed up and control reaction rates. Catalyst = Chemical agent that accelerates a reaction without being permanently changed in the process, so it can be used over and over.
Enzymes = Biological catalysts made of protein.
Before a reaction can occur, the reactants must absorb energy to break chemical bonds. This initial energy investment is the activation energy.
Free energy of activation (activation energy) = Amount of energy that reactant molecules must absorb to start a reaction (EA).
Transition state = Unstable condition of reactant molecules that have absorbed sufficient free energy to react.
Energy profile of an exergonic reaction:
1. Reactants must absorb enough
energy (EA) to reach the
transition state (uphill portion
of the curve). Usually the
absorption of thermal energy
from the surroundings is enough
to break chemical bonds.
2. Reaction occurs and energy is
released as new bonds form
(downhill portion of the curve).
3. G for the overall reaction is the
difference in free energy between
products and reactants. In an
exergonic reaction the free
energy of the products is less
than reactants.
Even though a reaction is energetically favorable, there must be an initial investment of activation energy (EA ).
The breakdown of biological macromolecules is exergonic. However, these molecules react very slowly at cellular temperatures because they cannot absorb enough thermal energy to reach transition state.
In order to make these molecules reactive when necessary, cells use biological catalysts called enzymes, which:
• Are proteins.
• Lower EA, so the transition state can be reached at cellular temperatures. • Do not change the nature of a reaction ( G), but only speed up a reaction that would have occurred anyway.
• Are very selective for which reaction they will catalyze.
B. Enzymes are substrate-specific
Enzymes are specific for a particular substrate, and that specificity depends upon the enzyme's three-dimensional shape.
Chapter 6 An Introduction to Metabolism 63
Substrate = The substance an enzyme acts on and makes more reactive. • An enzyme binds to its substrate and catalyzes its conversion to product. The enzyme is released in original form.
Substrate + enzyme enzyme-substrate complex product + enzyme • The substrate binds to the enzyme's active site.
Active site = Restricted region of an enzyme molecule which binds to the substrate. • Is usually a pocket or groove on the protein's surface.
• Formed with only a few of the enzyme's amino acids.
• Determines enzyme specificity which is based upon a compatible fit between the shape of an enzyme's active site and the shape of the substrate.
• Changes its shape in response to the substrate.
• As substrate binds to the active site, it induces the enzyme to change its shape.
• This brings its chemical groups into positions that enhance their ability to interact with the substrate and catalyze the reaction.
Induced fit = Change in the shape of an enzyme's active site, which is induced by the substrate (see Campbell, Figure 6.11).
C. The active site is an enzyme’s catalytic center
The entire enzymatic cycle is quite rapid (see Campbell, Figure 6.12). Steps in the catalytic cycle of enzymes:
1. Substrate binds to the active site forming an enzyme-substrate complex. Substrate is held in the active site by weak interactions (e.g., hydrogen bonds and ionic bonds).
2. Induced fit of the active site around the substrate. Side chains of a few amino acids in the active site catalyze the conversion of substrate to product. 3. Product departs active site and the enzyme emerges in its original form. Since enzymes are used over and over, they can be effective in very small amounts. Enzymes lower activation energy and speed up reactions by several mechanisms: • Active site can hold two or more reactants in the proper position so they may react.
• Induced fit of the enzyme's active site may distort the substrate's chemical bonds, so less thermal energy (lower G) is needed to break them during the reaction.
• Active site might provide a micro-environment conducive to a particular type of reaction (e.g., localized regions of low pH caused by acidic side chains on amino acids at the active site).
• Side chains of amino acids in the active site may participate directly in the reaction.
The initial substrate concentration partly determines the rate of an enzyme controlled reaction.
• The higher the substrate concentration, the faster the reaction - up to a limit. • If substrate concentration is high enough, the enzyme becomes saturated with substrate. (The active sites of all enzymes molecules are engaged.)
• When an enzyme is saturated, the reaction rate depends upon how fast the active sites can convert substrate to product.
• When enzyme is saturated, reaction rate may be increased by adding more enzyme.
64 Unit I The Chemistry of Life
D. A cell’s physical and chemical environment affects enzyme activity Each enzyme has optimal environmental conditions that favor the most active enzyme conformation.
1. Effects of temperature and pH
Optimal temperature allows the greatest number of molecular collisions without denaturing the enzyme.
• Enzyme reaction rate increases with increasing temperature. Kinetic energy of reactant molecules increases with rising temperature, which increases substrate collisions with active sites.
• Beyond the optimal temperature, reaction rate slows. The enzyme denatures when increased thermal agitation of molecules disrupts weak bonds that stabilize the active conformation.
• Optimal temperature range of most human enzymes is 35°– 40°C.
Optimal pH range for most enzymes is pH 6 – 8.
• Some enzymes operate best at more extremes of pH.
• For example, the digestive enzyme, pepsin, found in the acid environment of the stomach has an optimal pH of 2.
2. Cofactors
Cofactors = Small nonprotein molecules that are required for proper enzyme catalysis.
• May bind tightly to active site.
• May bind loosely to both active site and substrate.
• Some are inorganic (e.g., metal atoms of zinc, iron or copper).
• Some are organic and are called coenzymes (e.g., most vitamins).
3. Enzyme inhibitors
Certain chemicals can selectively inhibit enzyme activity (see Campbell, Figure 6.14).
• Inhibition may be irreversible if the inhibitor attaches by covalent bonds. • Inhibition may be reversible if the inhibitor attaches by weak bonds.
Competitive inhibitors = Chemicals that resemble an enzyme's normal substrate and compete with it for the active site.
• Block active site from the substrate.
• If reversible, the effect of these inhibitors can be overcome by increased substrate concentration.
Noncompetitive inhibitors = Enzyme inhibitors that do not enter the enzyme's active site, but bind to another part of the enzyme molecule.
• Causes enzyme to change its shape so the active site cannot bind substrate. • May act as metabolic poisons (e.g., DDT, many antibiotics).
• Selective enzyme inhibition is an essential mechanism in the cell for regulating metabolic reactions.
III. The Control of Metabolism
A. Metabolic pathways are regulated by controlling enzyme activity. Metabolic control often depends on allosteric regulation
1. Allosteric regulation
Allosteric site = Specific receptor site on some part of the enzyme molecule other than the active site.
Chapter 6 An Introduction to Metabolism 65
• Most enzymes with allosteric sites have two or more polypeptide chains, each with its own active site. Allosteric sites are often located where the subunits join.
• Allosteric enzymes have two conformations, one catalytically active and the other inactive (see Campbell, Figure 6.15) .
• Binding of an activator to an allosteric site stabilizes the active conformation.
• Binding of an inhibitor (noncompetitive inhibitor) to an allosteric site stabilizes the inactive conformation.
• Enzyme activity changes continually in response to changes in the relative proportions of activators and inhibitors (e.g., ATP/ADP).
• Subunits may interact so that a single activator or inhibitor at one allosteric site will affect the active sites of the other subunits.
2. Feedback inhibition
Feedback inhibition = Regulation of a metabolic pathway by its end product, which inhibits an enzyme within the pathway. For example:
Enzyme Enzyme Enzyme Enzyme Enzyme
1 2 3 4 5
threonine A B C D isoleucine
(initial substrate) (end product and allosteric inhibitor of enzyme 1)
Feedback Inhibition
Prevents the cell from wasting chemical resources by synthesizing more product than is necessary (see also Campbell, Figure 6.16).
3. Cooperativity
Substrate molecules themselves may enhance enzyme activity.
Cooperativity = The phenomenon where substrate binding to the active site of one subunit induces a conformational change that enhances substrate binding at the active sites of the other subunits (see Campbell, Figure 6.17).
B. The localization of enzymes within the cell helps order metabolism Cellular structure orders and compartmentalizes metabolic pathways (see Campbell, Figure 6.18).
• Some enzymes and enzyme complexes have fixed locations in the cell because they are incorporated into a membrane.
• Other enzymes and their substrates may be localized within membrane-enclosed eukaryotic organelles (e.g., chloroplasts and mitochondria).
REFERENCES
Atkins, P.W. The Second Law. New York, Oxford: W.H. Freeman and Company, 1984. A beautifully written, understandable description of the Second Law of Thermodynamics; addresses the role of the Second Law in life processes.
Campbell, N., et al. Biology. 5th ed. Menlo Park, California: Benjamin/Cummings, 1998. Lehninger, A.L., D.L. Nelson and M.M. Cox. Principles of Biochemistry. 2nd ed. New York: Worth, 1993.
CHAPTER 7
A TOUR OF THE CELL
OUTLINE
I. How We Study Cells
A. Microscopes provide windows to the world of the cell
B. Cell biologists can isolate organelles to study their functions
II. A Panoramic View of the Cell
A. Prokaryotic and eukaryotic cells differ in size and complexity
B. Internal membranes compartmentalize the functions of a eukaryotic cell III. The Nucleus and Ribosomes
A. The nucleus contains a eukaryotic cell’s genetic library
B. Ribosomes build a cell’s proteins
IV. The Endomembrane System
A. The endoplasmic reticulum manufactures membranes and performs many other biosynthetic functions
B. The Golgi apparatus finishes, sorts, and ships cell products
C. Lysosomes are digestive compartments
D. Vacuoles have diverse functions in cell maintenance
V. Other Membranous Organelles
A. Peroxisomes consume oxygen in various metabolic functions
B. Mitochondria and chloroplasts are the main energy transformers of cells VI. The Cytoskeleton
A. Provides structural support to cells for cell motility and regulation
VII. Cell Surfaces and Junctions
A. Plant cells are encased by cell walls
B. The extracellular matrix (ECM) of animal cells functions in support, adhesion, movement and development
C. Intercellular junctions help integrate cells into higher levels of structure and function
OBJECTIVES
After reading this chapter and attending lecture, the student should be able to: 1. Describe techniques used to study cell structure and function.
2. Distinguish between magnification and resolving power.
3. Describe the principles, advantages and limitations of the light microscope, transmission electron microscope and the scanning electron microscope.
4. Describe the major steps of cell fractionation and explain why it is a useful technique.
68 Unit II The Cell
5. Distinguish between prokaryotic and eukaryotic cells.
6. Explain why there are both upper and lower limits to cell size.
7. Explain why compartmentalization is important in eukaryotic cells. 8. Describe the structure and function of the nucleus, and briefly explain how the nucleus controls protein synthesis in the cytoplasm.
9. Describe the structure and function of a eukaryotic ribosome.
10. List the components of the endomembrane system, describe their structures and functions and summarize the relationships among them.
11. Explain how impaired lysosomal function causes the symptoms of storage diseases. 12. Describe the types of vacuoles and explain how their functions differ. 13. Explain the role of peroxisomes in eukaryotic cells.
14. Describe the structure of a mitochondrion and explain the importance of compartmentalization in mitochondrial function.
15. Distinguish among amyloplast, chromoplast and chloroplast.
16. Identify the three functional compartments of a chloroplast, and explain the importance of compartmentalization in chloroplast function.
17. Describe probable functions of the cytoskeleton.
18. Describe the structure, monomers and functions of microtubules, microfilaments and intermediate filaments.
19. Explain how the ultrastructure of cilia and flagella relates to their function. 20. Describe the development of plant cell walls.
21. Describe the structure and list some functions of the extracellular matrix in animal cells.
22. Describe the structure of intercellular junctions found in plant and animal cells, and relate their structure to function.
KEY TERMS
light microscope nucleolus thylakoid middle lamella resolving power ribosome granlakoids secondary cell wall organelle endomembrane system stroma extracellular matrix electron microscope endoplasmic reticulum (ER) cytoskeleton collagen TEM smooth ER microtubules proteoglycan SEM rough ER microfilaments fibronectin cell fractionation glycoprotein integrin intermediate filaments ultracentrifuges transport vesicles centrosome plasmodesmata cytoplasm Golgi apparatus centriole tight junctions prokaryotic cell phagocytosis flagella desmosomes nucleoid food vacuole cilia gap junctions eukaryotic cell contractile vacuole basal body
cytoplasm central vacuole dynein
cytosol peroxisome actin
plasma membrane mitochondria myosin
nucleus chloroplast pseudopodia
nuclear lamina cristae cytoplasmic streaming chromatin mitochondrial matrix cell wall
chromosome plastid primary cell wall
Chapter 7 A Tour of the Cell 69
LECTURE NOTES
All organisms are made of cells, the organism's basic unit of structure and function. The cell as a microcosm can be used to illustrate four themes integral to the text and course: 1. Theme of emergent properties. Life at the cellular level arises from interactions among cellular components.
2. Correlation of structure and function. Ordered cellular processes (e.g., protein synthesis, respiration, photosynthesis, cell-cell recognition, cellular movement, membrane production and secretion) are based upon ordered structures.
3. Interaction of organisms within their environment. Cells are excitable responding to environmental stimuli. In addition, cells are open systems that exchange materials and energy with their environment.
4. Unifying theme of evolution. Evolutionary adaptations are the basis for the correlation between structure and function.
I. How We Study Cells
A. Microscopes provide windows to the world of the cell
The microscope's invention and improvement in the seventeenth century led to the discovery and study of cells.
In 1665, Robert Hooke described cells using a light microscope. Modern light microscopy is based upon the same principles as microscopy first used by Renaissance scientists.
• Visible light is focused on a specimen with a condenser lens.
• Light passing through the specimen is refracted with an objective lens and an ocular lens. The specimen's image is thus magnified and inverted for the observer.
Two important concepts in microscopy are magnification and resolving power. • Magnification = How much larger an object is made to appear compared to its real size.
• Resolving power = Minimum distance between two points that can still be distinguished as two separate points.
• Resolution of a light microscope is limited by the wavelength of visible light. Maximum possible resolution of a light microscope is 0.2 μm.
• Highest magnification in a light microscope with maximum resolution is about 1000 times.
• By the early 1900s, optics in light microscopes were good enough to achieve the best resolution, so improvements since then have focused on improving contrast.
In the 1950s, researchers began to use the electron microscope which far surpassed the resolving power of the light microscope.
• Resolving power is inversely related to wavelength. Instead of light, electron microscopes use electron beams which have much shorter wavelengths than visible light.
• Modern electron microscopes have a practical resolving power of about 2 nm. • Enhanced resolution and magnification allowed researchers to clearly identify subcellular organelles and to study cell ultrastructure.
70 Unit II The Cell
• Two types of electron microscopes are the transmission electron microscope (TEM) and the scanning electron microscope.
The transmission electron microscope (TEM) aims an electron beam at a thin section of specimen which may be stained with metals to absorb electrons and enhance contrast.
• Electrons transmitted through the specimen are focused and the image magnified by using electromagnetic lenses (rather than glass lenses) to bend the trajectories of the charged electrons.
• Image is focused onto a viewing screen or film.
• Used to study internal cellular ultrastructure.
The scanning electron microscope (SEM) is useful for studying the surface of a specimen.
• Electron beam scans the surface of the specimen usually coated with a thin film of gold.
• Scanning beam excites secondary electrons on the sample's surface.
• Secondary electrons are collected and focused onto a viewing screen.
• SEM has a great depth of field and produces a three-dimensional image. Disadvantages of an electron microscope:
• Can usually only view dead cells because of the elaborate preparation required. • May introduce structural artifacts.
B. Cell biologists can isolate organelles to study their function
Modern cell biology integrates the study of cell structure (cytology) with the study of cell function. Cell fractionation is a technique that enables researchers to isolate organelles without destroying their function (see Campbell, Figure 7.3).
Cell fractionation = Technique which involves centrifuging disrupted cells at various speeds and durations to isolate components of different sizes, densities, and shapes. • Development of the ultracentrifuge made this technique possible.
• Ultracentrifuges can spin as fast as 80,000 rpm, applying a force of 500,000 g. The process of cell fractionation involves the following:
• Homogenization of tissue and its cells using pistons, blenders, or ultrasound devices.
• Centrifugation of the resulting homogenate at a slow speed. Nuclei and other larger particles settle at the bottom of the tube, forming a pellet.
• The unpelleted fluid or supernatant is decanted into another tube and centrifuged at a faster speed, separating out smaller organelles.
• The previous step is repeated, increasing the centrifugation speed each time to collect smaller and smaller cellular components from successive pellets.
• Once the cellular components are separated and identified, their particular metabolic functions can be determined.
Chapter 7 A Tour of the Cell 71
II. A Panoramic View of the Cell
A. Prokaryotic and eukaryotic cells differ in size and complexity Living organisms are made of either prokaryotic or eukaryotic cells—two major kinds of cells, which can be distinguished by structural organization.
Cytoplasm = Entire region between the nucleus and cell membrane
Cytosol = Semi-fluid medium found in the cytoplasm
1. Cell size
Size ranges of cells:
Range of cell size is limited by metabolic requirements. The lower limits are probably determined by the smallest size with enough:
• DNA to program metabolism.
• ribosomes, enzymes and cellular components to sustain life and reproduce. The upper limits of size are imposed by the surface area to volume ratio. As a cell increases in size, its volume grows proportionately more than its surface area (see Campbell, Figure 7.5).
• The surface area of the plasma membrane must be large enough for the cell volume, in order to provide an adequate exchange surface for oxygen, nutrients and wastes.
B. Internal membranes compartmentalize the functions of a eukaryotic cell The average eukaryotic cell has a thousand times the volume of the average prokaryotic cell, but only a hundred times the surface area. Eukaryotic cells compensate for the small surface area to volume ratio by having internal membranes which:
• Partition the cell into compartments.
• Have unique lipid and protein compositions depending upon their specific functions.
• May participate in metabolic reactions since many enzymes are incorporated directly into the membrane.
• Provide localized environmental conditions necessary for specific metabolic processes.
72 Unit II The Cell
• Sequester reactions, so they may occur without interference from incompatible metabolic processes elsewhere in the cell (see Campbell, Figure 7.6).
III. The Nucleus and Ribosomes
A. The nucleus contains a eukaryotic cell’s genetic library
Nucleus = A generally conspicuous membrane-bound cellular organelle in a eukaryotic cell; contains most of the genes that control the entire cell (see Campbell, Figure 7.9). • Averages about 5 μm diameter.
• Enclosed by a nuclear envelope.
Nuclear envelope = A double membrane which encloses the nucleus in a eukaryotic cell. • Is two lipid bilayer membranes separated by a space of about 20 to 40 nm. Each lipid bilayer has its own specific proteins.
• Attached to proteins on the envelope’s nuclear side is a network of protein filaments, the nuclear lamina, which stabilizes nuclear shape.
• Is perforated by pores (100 nm diameter), which are ordered by an octagonal array of protein granules.
• The envelope’s inner and outer membranes are fused at the lip of each pore.
• Pore complex regulates molecular traffic into and out of the nucleus.
• There is new evidence of an intranuclear framework of fibers, the nuclear matrix.
The nucleus contains most of the cell's DNA which is organized with proteins into a complex called chromatin.
Chromatin = Complex of DNA and histone proteins, which makes up chromosomes in eukaryotic cells; appears as a mass of stained material in nondividing cells. Chromosomes = Long threadlike association of genes, composed of chromatin and found in the nucleus of eukaryotic cells.
• Each species has a characteristic chromosome number.
• Human cells have 46 chromosomes, except egg and sperm cells, which have half or 23.
The most visible structure within the nondividing nucleus is the nucleolus. Nucleolus = Roughly spherical region in the nucleus of nondividing cells, which consists of nucleolar organizers and ribosomes in various stages of production.
• May be two or more per cell.
• Packages ribosomal subunits from:
• rRNA transcribed in the nucleolus.
• RNA produced elsewhere in the nucleus.
• Ribosomal proteins produced and imported from the cytoplasm.
• Ribosomal subunits pass through nuclear pores to the cytoplasm, where their assembly is completed.
Nucleolar organizers = Specialized regions of some chromosomes, with multiple copies of genes for rRNA (ribosomal RNA) synthesis.
The nucleus controls protein synthesis in the cytoplasm:
Chapter 7 A Tour of the Cell 73
Messenger RNA (mRNA) transcribed in the nucleus
from DNA instructions.
Passes through nuclear pores into cytoplasm.
Attaches to ribosomes where the genetic message
is translated into primary protein structure.
B. Ribosomes build a cell’s proteins
Ribosome = A cytoplasmic organelle that is the site for protein synthesis (see Campbell, Figure 7.10).
• Are complexes of RNA and protein
• Constructed in the nucleolus in eukaryotic cells
• Cells with high rates of protein synthesis have prominent nucleoli and many ribosomes (e.g., human liver cell has a few million).
Ribosomes function either free in the cytosol or bound to endoplasmic reticulum. Bound and free ribosomes are structurally identical and interchangeable.
Free ribosomes = Ribosomes suspended in the cytosol.
• Most proteins made by free ribosomes will function in the cytosol.
Bound ribosomes = Ribosomes attached to the outside of the endoplasmic reticulum. • Generally make proteins that are destined for membrane inclusion or export. • Cells specializing in protein secretion often have many bound ribosomes (e.g., pancreatic cells).
IV. The Endomembrane System
Biologists consider many membranes of the eukaryotic cell to be part of an endomembrane system.
• Membranes may be interrelated directly through physical contact.
• Membranes may be related indirectly through vesicles.
Vesicles = Membrane-enclosed sacs that are pinched off portions of membranes moving from the site of one membrane to another.
Membranes of the endomembrane system vary in structure and function, and the membranes themselves are dynamic structures changing in composition, thickness and behavior.
The endomembrane system includes:
• Nuclear envelope
• Endoplasmic reticulum
• Golgi apparatus
• Lysosomes
• Vacuoles
74 Unit II The Cell
• Plasma membrane (not actually an endomembrane, but related t o endomembrane system)
A. The endoplasmic reticulum manufactures membranes and performs many other biosynthetic functions
Endoplasmic reticulum (ER) = (Endoplasmic = within the cytoplasm; reticulum = network); extensive membranous network of tubules and sacs (cisternae) which sequesters its internal lumen (cisternal space) from the cytosol.
• Most extensive portion of endomembrane system.
• Continuous with the outer membrane of the nuclear envelope; therefore, the space between the membranes of the nuclear envelope is continuous with cisternal space.
There are two distinct regions of ER that differ in structure and function: smooth ER and rough ER (see Campbell, Figure 7.11).
1. Functions of smooth ER
Appears smooth in the electron microscope because its cytoplasmic surface lacks ribosomes. Smooth ER functions in diverse metabolic processes:
a. Participates in the synthesis of lipids, phospholipids and steroids • For example, vertebrate, particularly mammalian sex hormones and steroids secreted by the adrenal gland.
• Cells that produce and secrete these products are rich in smooth ER (e.g., testes, ovaries, skin oil glands).
b. Participates in carbohydrate metabolism
• Smooth ER in liver contains an embedded enzyme that catalyzes the final step in the conversion of glycogen to glucose (removes the phosphate from glucose-phosphate).
c. Detoxifies drugs and poisons
• Smooth ER, especially in the liver, contains enzymes which detoxify drugs and poisons.
• Enzymes catalyze the addition of hydroxyl groups to drugs and poisons. This makes them soluble in the cytosol, so they may be excreted from the body.
• Smooth ER in liver cells proliferates in response to barbiturates, alcohol and other drugs. This, in turn, may increase drug tolerance.
d. Stores calcium ions necessary for muscle contraction
• In a muscle cell, the ER membrane pumps Ca++ from the cytosol into the cisternal space.
• In response to a nerve impulse, Ca++ leaks from the ER back into the cytosol, which triggers muscle cell contraction.
2. Rough ER and protein synthesis
Rough ER:
• Appears rough under an electron microscope because the cytoplasmic side is studded with ribosomes.
• Is continuous with outer membrane of the nuclear envelope (which may also be studded with ribosomes on the cytoplasmic side).
• Manufactures secretory proteins and membrane.
Chapter 7 A Tour of the Cell 75
Proteins destined for secretion are synthesized by ribosomes attached to rough ER:
Ribosomes attached to rough ER synthesize
secretory proteins.
Growing polypeptide is threaded through ER
membrane into the lumen or cisternal space.
Protein folds into its native conformation.
If destined to be a glycoprotein, enzymes
localized in the ER membrane catalyze the
covalent bonding of an oligosaccharide to the
secretory protein.
Protein departs in a transport vesicle pinched off
from transitional ER adjacent to the rough ER site of
production.
Glycoprotein = Protein covalently bonded to carbohydrate.
Oligosaccharide = Small polymer of sugar units.
Transport vesicle = Membrane vesicle in transit from one part of the cell to another.
3. Rough ER and membrane production
Membranes of rough ER grow in place as newly formed proteins and phospholipids are assembled:
• Membrane proteins are produced by ribosomes. As a polypeptide grows, it is inserted directly into the rough ER membrane where it is anchored by hydrophobic regions of the proteins.
• Enzymes within the ER membrane synthesize phospholipids from raw materials in the cytosol.
• Newly expanded ER membrane can be transported as a vesicle to other parts of the cell.
76 Unit II The Cell
B. The Golgi apparatus finishes, sorts, and ships cell products
Many transport vesicles leave the ER and travel to the Golgi apparatus. Golgi apparatus = Organelle made of stacked, flattened membranous sacs (cisternae), that modifies, stores and routes products of the endoplasmic reticulum (see Campbell, Figure 7.12).
cis face
(forming face)
Transport vesicle
from ER
Vesicle forming
Free Transport
vesicle from
the Golgi
Trans face
(maturing face)
Plasma
• Membranes of the cisternae sequester cisternal space from the cytosol. • Vesicles may transport macromolecules between the Golgi and other cellular structures.
• Has a distinct polarity. Membranes of cisternae at opposite ends differ in thickness and composition.
• Two poles are called the cis face (forming face) and the trans face (maturing face).
• Cis face, which is closely associated with transitional ER, receives products by accepting transport vesicles from the ER. A vesicle fuses its membrane to the cis face of the Golgi and empties its soluble contents into the Golgi's cisternal space.
• Trans face pinches off vesicles from the Golgi and transports molecules t o other sites.
Enzymes in the Golgi modify products of the ER in stages as they move through the Golgi stack from the cis to the trans face:
• Each cisternae between the cis and trans face contains unique combinations of enzymes.
• Golgi products in transit from one cisternae to the next, are carried in transport vesicles.
During this process, the Golgi:
• Alters some membrane phospholipids.
• Modifies the oligosaccharide portion of glycoproteins.
• Manufactures certain macromolecules itself (e.g., hyaluronic acid). • Targets products for various parts of the cell.
• Phosphate groups or oligosaccharides may be added to Golgi products as molecular identification tags.
• Membranous vesicles budded from the Golgi may have external molecules that recognize docking sites on the surface of certain other organelles. • Sorts products for secretion. Products destined for secretion leave the trans face in vesicles which eventually fuse with the plasma membrane.
Chapter 7 A Tour of the Cell 77
C. Lysosomes are digestive compartments
Lysosome = An organelle which is a membrane-enclosed bag of hydrolytic enzymes that digest all major classes of macromolecules (see Campbell, Figure 7.13). • Enzymes include lipases, carbohydrases, proteases, and nucleases.
• Optimal pH for lysosomal enzymes is about pH 5.
• Lysosomal membrane performs two important functions:
• Sequesters potentially destructive hydrolytic enzymes from the cytosol.
• Maintains the optimal acidic environment for enzyme activity by pumping H+s inward from the cytosol to the lumen.
• Hydrolytic enzymes and lysosomal membrane are synthesized in the rough ER and processed further in the Golgi apparatus.
• Lysosomes probably pinch off from the trans face of the Golgi apparatus (see Campbell, Figure 7.14).
1. Functions of lysosomes
a. Intracellular digestion
Phagocytosis = (Phago = to eat; cyte = cell); cellular process of ingestion, in which the plasma membrane engulfs particulate substances and pinches off to form a particle-containing vacuole.
• Lysosomes may fuse with food-filled vacuoles, and their hydrolytic enzymes digest the food.
• Examples are Amoeba and other protists which eat smaller organisms or food particles.
• Human cells called macrophages phagocytize bacteria and other invaders.
b. Recycle cell's own organic material
• Lysosomes may engulf other cellular organelles or part of the cytosol and digest them with hydrolytic enzymes (autophagy).
• Resulting monomers are released into the cytosol where they can be recycled into new macromolecules.
c. Programmed cell destruction
Destruction of cells by their own lysosomes is important during metamorphosis and development.
2. Lysosomes and human disease
Symptoms of inherited storage diseases result from impaired lysosomal function. Lack of a specific lysosomal enzyme causes substrate accumulation which interferes with lysosomal metabolism and other cellular functions.
• In Pompe's disease, the missing enzyme is a carbohydrase that breaks down glycogen. The resulting glycogen accumulation damages the liver.
• Lysosomal lipase is missing or inactive in Tay-Sachs disease, which causes lipid accumulation in the brain.
D. Vacuoles have diverse functions in cell maintenance
Vacuole = Organelle which is a membrane-enclosed sac that is larger than a vesicle (transport vesicle, lysosome, or microbody).
Vacuole types and functions:
Food vacuole = Vacuole formed by phagocytosis which is the site of intracellular digestion in some protists and macrophages (see Campbell, Figure 7.14). Contractile vacuole = Vacuole that pumps excess water from the cell; found in some freshwater protozoa.
78 Unit II The Cell
Central vacuole = Large vacuole found in most mature plant cells (see Campbell, Figure 7.15)
• Is enclosed by a membrane called the tonoplast which is part of the endomembrane system
• Develops by the coalescence of smaller vacuoles derived from the ER and Golgi apparatus
• Is a versatile compartment with many functions:
• Stores organic compounds (e.g., protein storage in seeds)
• Stores inorganic ions (e.g., K+ and Cl-)
• Sequesters dangerous metabolic by-products from the cytoplasm
• Contains soluble pigments in some cells (e.g., red and blue pigments in flowers)
• May protect the plant from predators by containing poisonous or unpalatable compounds
• Plays a role in plant growth by absorbing water and elongating the cell • Contributes to the large ratio of membrane surface area to cytoplasmic volume. (There is only a thin layer of cytoplasm between the tonoplast
and plasma membrane.)
E. A summary of relationships among endomembranes
Components of the endomembrane system are related through direct contact or through vesicles (see Campbell, Figure 7.16).
Nuclear Envelope is an extension of Rough ER is confluent with Smooth ER
membrane and secretory
proteins produced in
ER are transported in
Vesicles
fuse with the
forming face of
Golgi Apparatus
pinches off
maturing face
Vesicles
fuse with and add to
give rise plasma membrane to and may release cellular products
to outside
Lysosomes Plasma
and Vacuoles Membrane
V. Other Membranous Organelles
A. Peroxisomes consume oxygen in various metabolic functions
Peroxisomes = Membrane-bound organelles that contain specialized teams of enzymes for specific metabolic pathways; all contain peroxide-producing oxidases.
Chapter 7 A Tour of the Cell 79
• Bound by a single membrane
• Found in nearly all eukaryotic cells
• Often have a granular or crystalline core which is a dense collection of enzymes (see Campbell, Figure 7.17)
• Contain peroxide-producing oxidases that transfer hydrogen from various substrates to oxygen, producing hydrogen peroxide
RH2 + O2oxidase R + H2O2
• Contain catalase, an enzyme that converts toxic hydrogen peroxide to water 2H2O2catalase 2H2O + O2
• Peroxisomal reactions have many functions, some of which are:
• Breakdown of fatty acids into smaller molecules (acetyl CoA). The products are carried to the mitochondria as fuel for cellular respiration.
• Detoxification of alcohol and other harmful compounds. In the liver, peroxisomes enzymatically transfer H from poisons to O2.
• Specialized peroxisomes (glyoxysomes) are found in heterotrophic fat-storing tissue of germinating seeds.
• Contain enzymes that convert lipid to carbohydrate.
• These biochemical pathways make energy stored in seed oils available for the germinating seedling.
• Current thought is that peroxisome biogenesis occurs by pinching off from preexisting peroxisomes. Necessary lipids and enzymes are imported from the cytosol.
B. Mitochondria and chloroplasts are the main energy transformers of cells Mitochondria and chloroplasts are organelles that transduce energy acquired from the surroundings into forms useable for cellular work.
• Enclosed by double membranes (see Campbell, Figure 7.18).
• Membranes are not part of endomembrane system. Rather than being made in the ER, their membrane proteins are synthesized by free ribosomes in the cytosol and by ribosomes located within these organelles themselves.
• Contain ribosomes and some DNA that programs a small portion of their own protein synthesis, though most of their proteins are synthesized in the cytosol programmed by nuclear DNA.
• Are semiautonomous organelles that grow and reproduce within the cell.
1. Mitochondria
Mitochondria = Organelles which are the sites of cellular respiration, a catabolic oxygen-requiring process that uses energy extracted from organic macromolecules to produce ATP.
• Found in nearly all eukaryotic cells
• Number of mitochondria per cell varies and directly correlates with the cell's metabolic activity
• Are about 1 mm in diameter and 1-10 mm in length
• Are dynamic structures that move, change their shape and divide
80 Unit II The Cell
Structure of the mitochondrion:
• Enclosed by two membranes that have their own unique combination of proteins embedded in phospholipid bilayers (see Campbell, Figure 7.18)
• Smooth outer membrane is highly permeable to small solutes, but it blocks passage of proteins and other macromolecules
• Convoluted inner membrane contains embedded enzymes that are involved in cellular respiration. The membrane's many infoldings or cristae increase the surface area available for these reactions to occur.
• The inner and outer membranes divide the mitochondrion into two internal compartments:
a. Intermembrane space
• Narrow region between the inner and outer mitochondrial membranes.
• Reflects the solute
composition of the cytosol,
because the outer membrane
is permeable to small solute
molecules.
b. Mitochondrial matrix
Intermembrane space
Mitochondrial
matrix
Outer membrane Cristae
• Compartment enclosed by the inner mitochondrial membrane
• Contains enzymes that catalyze many metabolic steps of cellular respiration
• Some enzymes of respiration and ATP production are actually embedded in the inner membrane.
2. Chloroplasts
Plastids = A group of plant and algal membrane-bound organelles that include amyloplasts, chromoplasts and chloroplasts.
Amyloplasts = (Amylo = starch); colorless plastids that store starch; found in roots and tubers.
Chromoplasts = (Chromo = color); plastids containing pigments other than chlorophyll; responsible for the color of fruits, flowers and autumn leaves.
Chloroplasts = (Chloro = green); chlorophyll-containing plastids which are the sites of photosynthesis.
• Found in eukaryotic algae, leaves and other green plant organs. • Are lens-shaped and measure about 2 mm by 5 mm.
• Are dynamic structures that change shape, move and divide.
Structure of the chloroplast:
Chloroplasts are divided into three functional compartments by a system of membranes (see also Campbell, Figure 7.19):
a. Intermembrane space
The chloroplast is bound by a double membrane which partitions its contents from the cytosol. A narrow intermembrane space separates the two membranes.
Chapter 7 A Tour of the Cell 81
b. Thylakoid space
Thylakoids form another membranous system within the chloroplast. The thylakoid membrane segregates the interior of the chloroplast into two compartments: thylakoid space and stroma.
Intermembrane space
Stroma
Thylakoid space
• Thylakoid space = Space inside the thylakoid
• Thylakoids = Flattened membranous sacs inside the chloroplast
• Chlorophyll is found in the thylakoid membranes.
• Thylakoids function in the steps of photosynthesis that initially convert light energy to chemical energy.
• Some thylakoids are stacked into grana.
Grana = (Singular, granum); stacks of thylakoids in a chloroplast.
c. Stroma
Photosynthetic reactions that use chemical energy to convert carbon dioxide to sugar occur in the stroma.
Stroma = Viscous fluid outside the thylakoids
VI. The Cytoskeleton
A. Provides structural support to the cells for cell motility and regulation It was originally thought that organelles were suspended in a formless cytosol. Technological advances in both light and electron microscopy (e.g., high voltage E.M.) revealed a three-dimensional view of the cell, which showed a network of fibers throughout the cytoplasm—the cytoskeleton. The cytoskeleton plays a major role in organizing the structures and activities of the cell.
Cytoskeleton = A network of fibers throughout the cytoplasm that forms a dynamic framework for support and movement and regulation (see Campbell, Figure 7.20). • Gives mechanical support to the cell and helps maintain its shape
• Enables a cell to change shape in an adaptive manner
• Associated with motility by interacting with specialized proteins called motor molecules (e.g., organelle movement, muscle contraction, and locomotor organelles)
• Play a regulatory role by mechanically transmitting signals from cell’s surface to its interior
• Constructed from at least three types of fibers: microtubules (thickest), microfilaments (thinnest), and intermediate filaments (intermediate in diameter) (see Campbell, Table 7.2)
1. Microtubules
Found in cytoplasm of all eukaryotic cells, microtubules:
• Are straight hollow fibers about 25 nm in
diameter and 200 nm – 25 μm in length
• Are constructed from globular proteins
called tubulin that consists of one -
tubulin and one -tubulin molecule
82 Unit II The Cell
• Begin as two-dimensional sheets of tubulin units, which roll into tubes • Elongate by adding tubulin units to its ends
• May be disassembled and the tubulin units recycled to build microtubules elsewhere in the cell
Functions of microtubules include:
• Cellular support; these microtubule function as compression-resistant girders to reinforce cell shape
• Tracks for organelle movement (see Campbell, Figure 7.21). Protein motor molecules (e.g., kinesin) interact with microtubules to translocate organelles (e.g., vesicles from the Golgi to the plasma membrane).
• Separation of chromosomes during cell division
a. Centrosomes and centrioles
Centriole = Pair of cylindrical structures located in the centrosome of in animal cells, composed of nine sets of triplet microtubules arranged in a ring (see Campbell, Figure 7.22).
• Are about 150 nm in diameter and are arranged at right angles to each other.
• Pair of centrioles located within the centrosome, replicate during cell division.
• May organize microtubule assembly during cell division, but must not be mandatory for this function since plants lack centrioles.
b. Cilia and flagella
Cilia and flagella = Locomotor organelles found in eukaryotes that are formed from a specialized arrangement of microtubules.
• Many unicellular eukaryotic organisms are propelled through the water by cilia or flagella and motile sperm cells (animals, algae, some plants)
are flagellated.
• May function to draw fluid across the surface of stationary cells (e.g., ciliated cells lining trachea).
Chapter 7 A Tour of the Cell 83
Ultrastructure of cilia and flagella:
• Are extensions of plasma membrane with a core of microtubules (see Campbell, Figure 7.24)
• Microtubular core is made
of nine doublets of microtubules arranged in a ring with two single microtubules in the center (9 + 2 pattern).
Outer microtubule doublet
Dynein side
arms
Central
microtubule
• Each doublet is a pair of attached microtubules. One of the pair shares a portion of the other's wall.
• Each doublet is connected to the center of the ring
Protein spoke
by radial spokes that end near the central microtubules.
9+2 Pattern in Cross Section
• Each doublet is attached to the neighboring doublet by a pair of side arms. Many pairs of side arms are evenly spaced along the doublet's length.
• Structurally identical to centrioles, basal bodies anchor the microtubular assemblies.
Basal body = A cellular structure, identical to a centriole, that anchors the microtubular assembly of cilia and flagella.
• Can convert into a centriole and vice versa
• May be a template for ordering tubulin into the microtubules of newly forming cilia or flagella. As cilia and flagella continue to grow, new tubulin subunits are added to the tips, rather than to the bases.
The unique ultrastructure of cilia and flagella is necessary for them to function: • Sidearms are made of dynein, a large protein motor molecule that changes its conformation in the presence of ATP as an energy source. • A complex cycle of movements caused by dynein's conformational changes, makes the cilium or flagellum bend (see Campbell, Figure 7.25):
• In cilia and flagella, linear displacement of dynein sidearms is translated into a bending by the resistance of the radial spokes. Working against this resistance, the "dynein-walking" distorts the microtubules, causing them to bend.

Comments
Post a Comment